Fig. 20.1
Cartoon of CSF biomarkers and their relation to physiological and pathogenic processes. Aβ amyloid β, BACE1 β-site APP cleaving enzyme 1, sAPP soluble amyloid precursor protein
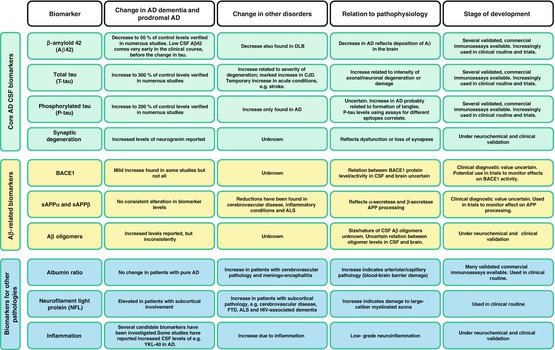
Fig. 20.2
Table of CSF biomarkers reviewed in this chapter. AD Alzheimer’s disease, ALS amyotrophic lateral sclerosis, BACE1 β-site APP cleaving enzyme 1, CJD Creutzfeldt-Jakob disease, CSF cerebrospinal fluid, DLB dementia with Lewy bodies, FTD frontotemporal dementia, HIV human immunodeficiency virus, sAPP soluble amyloid precursor protein
20.2 Core CSF AD Biomarkers
The CSF biomarkers for AD that have been most successful reflect the core cerebral pathologies of the disease.
20.2.1 Aβ42
Aβ is produced during normal cell metabolism and is secreted into the CSF (Seubert et al. 1992). The finding that Aβ42 is the most common isoform of Aβ in senile plaques led to development of assays for its quantification (Jarrett et al. 1993). The levels of CSF Aβ1-42 (Aβ42) are approximately 50 % lower in AD patients compared to controls, which has been shown with several different enzyme-linked immunosorbent assay (ELISA) methods (Sunderland et al. 2003; Blennow 2004). The most prevalent explanation for the reduced Aβ42 in CSF is that the sequestration into plaques in the brain leaves less available for diffusion into the CSF. Cerebral plaque load at autopsy correlates inversely with Aβ42 levels in lumbar CSF antemortem and ventricular CSF postmortem (Tapiola et al. 2009; Strozyk et al. 2003). Studies using PET for in vivo amyloid imaging have also found an inverse relationship between amyloid brain load and CSF Aβ42 (Fagan et al. 2006; Forsberg et al. 2008; Tolboom et al. 2009; Grimmer et al. 2009). Similar results have been obtained using the PET ligand 18FFDDNP, which is believed to bind to both plaques and tangles (Tolboom et al. 2009). These findings provide support of the notion that CSF Aβ42 is a measure of fibrillar Aβ42 and cerebral plaque load. Importantly, recent large studies have shown that the agreement between CSF Aβ42 and amyloid PET measurements is very high, with more than 90 % concordance, demonstrating that these methods give similar diagnostic information (Palmqvist et al. 2014).
20.2.2 P-tau
The amount of P-tau in CSF is commonly measured by ELISAs targeting tau phosphorylated at either threonine 181 or threonine 231 (Kohnken et al. 2000; Vanmechelen et al. 2000). These assays correlate well and show a similar association with AD (Hampel et al. 2004). Patients with AD have increased levels of CSF P-tau, in most studies around 200–300 % of control levels, and the levels in CSF have been shown to correlate with cerebral tangle pathology at autopsy (Blennow 2004; Tapiola et al. 2009; Buerger et al. 2006). In a study where cortical biopsies were taken from living patients with normal pressure hydrocephalus, a correlation between the amount of hyperphosphorylated tau in the biopsies and P-tau levels in CSF was found (Seppala et al. 2012). This supports the hypothesis that CSF P-tau reflects cerebral tangle pathology and the phosphorylation state of tau.
20.2.3 T-tau
CSF total tau corresponds to the intensity of neuronal and axonal damage and degeneration, which has been demonstrated in several studies. Levels increase after cardiac arrest, stroke, and brain trauma and are associated with amount of damaged tissue as well as clinical outcome (Hesse et al. 2001; Ost et al. 2006; Zetterberg et al. 2006; Rosen et al. 2014). In patients with Creutzfeldt-Jakob disease (CJD), a rapid rate of neurodegeneration is present, which is reflected in CSF by very high levels of T-tau (Otto et al. 1997). There are several isoforms and phosphorylation sites of the tau protein (Portelius et al. 2008). The ELISA assay most frequently used is capable of detecting all isoforms independent of phosphorylation state (Blennow et al. 1995). Studies have consistently shown that patients with AD have increased levels with about 300 % compared with normal controls (Sunderland et al. 2003; Blennow 2004). Also, it has been shown that levels of CSF T-tau antemortem correlate with the load of tangle pathology at autopsy, which indicates that release of tau from tangle-bearing neurons may contribute to levels of T-tau. In line with this, binding of the PET ligand 18FFDDNP has been shown to correlate with CSF T-tau levels (Tolboom et al. 2009).
20.3 Core Biomarkers in Various Stages of AD
20.3.1 Preclinical AD
Studies on preclinical AD have either been cross sectional, including patients with FAD, or longitudinal, with healthy elderly cohorts followed over time. The cross-sectional studies have calculated expected age of disease onset, which they relate biomarker levels to. One study showed that mutation carriers had elevated levels of CSF T-tau 15 years before expected symptom onset and reduced levels of CSF Aβ42 10 years before expected symptom onset (Bateman et al. 2012). Another study showed increased CSF Aβ42 in mutation carriers 20 years before estimated MCI onset (Reiman et al. 2012). Other studies that included cognitively normal mutation carriers closer to expected onset of AD have found increased levels of P-tau or T-tau, reduced Aβ42, and reduced Aβ42: Aβ40 ratio in CSF in these individuals compared with controls (Ringman et al. 2012; Moonis et al. 2005; Ringman et al. 2008).
The longitudinal studies have included cognitively healthy individuals, from which baseline biomarker levels have been related to future decrease in cognitive function or development of MCI or AD. CSF levels of Aβ42 alone or in combination with T-tau or P-tau have been associated with impending cognitive impairment (Gustafson et al. 2007; Fagan et al. 2007; Stomrud et al. 2007). Increased rates of brain atrophy have been found in cognitively normal individuals with low levels of CSF Aβ42 (Schott et al. 2010; Fagan et al. 2009; Fjell et al. 2010). Increased ratio of T-tau:Aβ42 or low levels of CSF Aβ42 have been found to predict conversion to MCI in cognitively normal individuals or AD in nondemented elderly individuals, respectively (Li et al. 2007; Skoog et al. 2003). One study found that the predictive ability of CSF Aβ42 for the development of MCI or AD in individuals with subjective complaints was superior to that of P-tau or T-tau (van Harten et al. 2012). However, long-term prospective CSF studies on cognitively normal individuals followed for 10–20 years until development of AD are missing. Also, the longitudinal studies are in fact cross sectional with respect to biomarker measurements, since baseline biomarker data has been correlated with longitudinal clinical outcome. True longitudinal studies with repeated biomarker measurements to determine the longitudinal biomarker trajectories are needed.
20.3.2 MCI Due to AD
Patients with MCI have a risk for AD over a 4.5-year period that is roughly tripled compared to cognitively normal controls (Bennett et al. 2002). The yearly progression rate from MCI to AD is around 10–15 % (Petersen 2004). Hence, long follow-up periods are required so that late converters are caught. Also, it is important to remember the heterogeneous nature of MCI, since affected individuals may progress to other dementias than AD, including vascular dementia (VaD), frontotemporal dementia (FTD), and dementia with Lewy bodies (DLB), or remain relatively stable in cognition (Petersen 2003). Patients with impending AD can be accurately distinguished from stable MCI patients by the CSF biomarker profile of low levels of Aβ42 in combination with high levels of P-tau and T-tau (Blennow and Hampel 2003). Different combinations of these biomarkers have been used in several studies to successfully predict AD in MCI patients (Hansson et al. 2006; Buchhave et al. 2012; Hertze et al. 2010; Visser et al. 2009; Shaw et al. 2009; Mattsson et al. 2009; Johansson et al. 2011a). CSF T-tau and P-tau levels have been associated with brain atrophy in early stage MCI/AD (Fagan et al. 2009). Also, the levels of these two biomarkers were higher in MCI patients with a more rapid progression to AD, indication that they may be related to disease progress (Buchhave et al. 2012).
20.3.3 AD with Dementia
The combination of Aβ42, P-tau, and T-tau can accurately distinguish AD patients from controls, with sensitivity and specificity over 80 % (Blennow 2004; Blennow and Hampel 2003). P-tau is the most AD specific of the biomarkers and may therefore be valuable when differentiating against other dementias (Hampel et al. 2010). Increased levels of P-tau have been found in patients with AD compared with patients suffering from FTD and VaD, DLB, and Parkinson disease with dementia (Hampel et al. 2004; Hall et al. 2012a). However, a large study on patients with AD as well as DLB, FTD, and VaD showed that an AD biomarker profile was present in a substantial part of the non-AD patients (Schoonenboom et al. 2012). Potential reasons for this overlap may be misdiagnosis or the presence of mixed dementias. Future autopsy studies will hopefully shed light on this issue. The presence of mixed pathologies increases with age, but the applicability of the core AD CSF biomarkers has been assessed in AD patients in different ages (Mattsson et al. 2012a). It was found that the accuracy decreased with age, but the combination of the biomarkers gave sufficiently strong results for the biomarkers to be used in old patients. Some studies have evaluated the longitudinal alterations of the core biomarkers in AD patients. Levels of Aβ42 and P-tau remain largely unaltered over time, while data concerning T-tau is more inconclusive (Blennow et al. 2007; Zetterberg et al. 2007; Mattsson et al. 2012b; Buchhave et al. 2009; Le Bastard et al. 2013). Some studies have found an increase of T-tau over time, while others have found stable levels. However, high levels of P-tau and T-tau have been associated with a more rapid disease progression (Samgard et al. 2010; Wallin et al. 2010; van Rossum et al. 2012).
20.3.4 Autopsy-Verified AD
The diagnostic performance of CSF AD biomarkers has been evaluated by performing subsequent autopsy of patients after death. They can with high sensitivity and specificity discriminate AD from cognitively normal elderly as well as patients with other dementias, such as FTD, DLB, and VaD (Sunderland et al. 2003; Shaw et al. 2009; Clark et al. 2003; Bian et al. 2008; Koopman et al. 2009). CSF biomarkers provide similar or better diagnostic accuracy compared with only using clinical criteria for diagnosis.
20.4 Aβ-Related Biomarkers
20.4.1 BACE1
The activity of BACE1 has been assessed in AD patients, but the results have not been consistent. Smaller studies have found increased BACE1 activity in CSF from AD patients (Holsinger et al. 2004; Holsinger et al. 2006; Verheijen et al. 2006). One study found that the activity was increased in patients with MCI and dementia due to AD compared with controls (Zetterberg et al. 2008). Another study found elevated levels in MCI patients, but not in AD patients (Mulder et al. 2010; Zhong et al. 2007). Finally, three studies found no differences between AD patients and controls (Mulder et al. 2010; Zhong et al. 2007; Rosén et al. 2012; Perneczky et al. 2014). However, in one of these studies, AD patients were stratified according to disease severity, which revealed that patients with moderate–severe dementia had lower levels of BACE1 activity than patients with mild dementia (Rosén et al. 2012). Although there is no clear-cut role for BACE1 activity in the diagnosis of AD, it may be useful in clinical trials on BACE1-inhibitors to monitor target engagement of the drug.
20.4.2 sAPPα/sAPPβ
The soluble ectodomain of APP (sAPP) is released when the protein is cut by α- or β-secretase. The levels of sAPPα and sAPPβ correlate very well in CSF in both AD patients and controls (Zetterberg et al. 2008). A large number of studies have not shown any differences in levels when comparing AD patients with controls (Hertze et al. 2010; Johansson et al. 2011a; Zetterberg et al. 2008; Perneczky et al. 2014; Rosen et al. 2012; Olsson et al. 2003). Some studies report elevated levels of sAPPβ in MCI patients compared to controls or in MCI patients with impending AD (Olsson et al. 2003; Perneczky et al. 2011). The latter differences were not seen in another study (Hertze et al. 2010). Studies using the core CSF AD biomarkers to characterize patients have found elevated levels of sAPPα and sAPPβ in patients with MCI or dementia that had a biomarker profile indicative of AD pathology, but with large overlaps between diagnostic groups (Lewczuk et al. 2010, 2012; Gabelle et al. 2010). The diagnostic value of sAPP levels appears low, but they may be utilized in clinical trials for studying effects on the APP metabolism, where a dose-dependent decrease in CSF sAPPβ indicates target engagement in clinical trials on BACE1 inhibitors (May et al. 2011).
20.4.3 Aβ Oligomers
The association between brain amyloid burden and neuronal loss is poor (Gomez-Isla et al. 1997). However, it has been shown that soluble oligomers of Aβ are capable of inhibiting long-term potentiation in vivo and cause abnormal tau phosphorylation and neuritic dystrophy (Walsh et al. 2002; Zempel et al. 2010; Jin et al. 2011; De Felice et al. 2008). Hence, many studies report attempts to measure the CSF levels of Aβ oligomers in patients with AD. This has proven to be difficult and studies have shown that the CSF levels of Aβ oligomers are very low, probably less than 1 % of total Aβ levels, which make reliable quantification challenging. Elevated oligomer levels have been found in the brains and CSF of AD patients (Bruggink et al. 2013; Shankar et al. 2008; Gao et al. 2010; Fukumoto et al. 2010; Georganopoulou et al. 2005; Pitschke et al. 1998). Increased levels were also reported in cognitively normal subjects with a CSF biomarker profile that indicated AD neuropathology (Handoko et al. 2013). Other studies have not been able to find altered levels of Aβ oligomers in CSF from AD patients (Bruggink and Jongbloed 2012; Santos et al. 2012; Yang et al. 2013). Apart from the difficulties of measuring minute amounts of Aβ oligomers in CSF samples, it is also possible that different assays measure different variants of Aβ oligomers, which might explain the divergent results. The studies provide little characterization of what the various assays are measuring.
20.5 Biomarkers for Other Pathologies
20.5.1 Blood-Brain Barrier Disturbances
The major biomarker for the integrity of the blood-brain barrier is the ratio between albumin measured in CSF and blood. This albumin ratio is typically normal in AD patients but tends to be elevated in patients with cerebral small vessel disease (Blennow et al. 1990; Wallin et al. 1999). An increased ratio may also be found in Lyme disease (neuroborreliosis), together with findings such as monocytosis and signs of immunoglobulin production in the CSF (Tumani et al. 1995). New potential biomarkers of the blood-brain barrier that need further verification include secretory Ca2+-dependent phospholipase A2 activity and antithrombin III (Chalbot et al. 2010; Zetterberg et al. 2009).
20.5.2 Neurofilament Light (NFL) Protein
NFL is mainly present in large-caliber myelinated axons, and increased levels are seen in conditions with subcortical axonal degeneration, such as cerebral small vessel disease, FTD, and human immunodeficiency virus (HIV)-associated dementia (Rosengren et al. 1999; Agren-Wilsson et al. 2007; Wallin and Sjogren 2001; de Jong et al. 2007; Landqvist Waldo et al. 2013; Gisslen et al. 2007). It may thus be of value in the differential diagnostics of AD. The simultaneous finding of elevated T-tau and NFL levels in CSF indicates mixed forms of AD and cerebrovascular disease, which appears to be very common in unselected patients undergoing evaluation because of a suspected neurodegenerative disease (Skillback et al. 2013).
20.5.3 Inflammatory, Glial, and Oxidative Stress Biomarkers
Neuroinflammation is a nonspecific feature of AD, and microglial cells can be found around senile plaques. In the 1990s and early 2000s, several studies reporting alterations in different inflammatory markers in CSF of AD patients (e.g., α1-antichymotrypsin, isoprostane, the interleukins, tumor necrosis factor α, interferon-gamma, complement C1q, and transforming growth factor β) were published (Craig-Schapiro et al. 2009; Zetterberg et al. 2004). The results have though been very inconsistent. A meta-analysis of cytokines in AD in 2010 showed great diversity in the study results (Swardfager et al. 2010). Reasons for discrepancies could include methodological differences (e.g., in the procedures for CSF collection and processing, assay differences, and criteria used for subject ascertainment), prevalence of comorbidities in the studied cohorts, and methods of diagnosis.
A recently described mutation in the microglia-controlling triggering receptor expressed on myeloid cells-2 (TREM2) gene was associated with an increased risk of AD. This has increased the interest of the role of microglia in the development of AD. Microglia are the macrophages of the brain. The enzyme chitotriosidase is secreted by activated macrophages and its levels of which are increased in the lysosomal storage disorder Gaucher disease (Renkema et al. 1998; Hollak et al. 1994). Studies on AD patients have revealed increased levels in CSF, although with overlaps with control groups (Mattsson et al. 2011a; Watabe-Rudolph et al. 2012; Rosén et al. 2014). A glycoprotein that has great homology with chitotriosidase but lacks its enzymatic activity is YKL-40 (Hakala et al. 1993). It is present in both microglial cells and astrocytes, and increased levels have been found in AD patients compared with controls (Rosén et al. 2014; Craig-Schapiro et al. 2010; Perrin et al. 2011; Olsson et al. 2013). One study did not find altered levels in AD patients compared with controls (Mattsson et al. 2011a).
A degree of oxidative stress is present in brains of AD patients and contributes to neuronal damage. Free radicals can cause lipid peroxidation, a feature that may be studied using biomarkers. A compound named isoprostane is produced by free radical-dependent peroxidation of arachidonic acid (Morrow and Roberts 1997). A certain subset, called F2-isoprostane, has been found in increased levels in CSF from AD patients (Montine et al. 1999; Pratico et al. 2000; Montine et al. 2001; Montine et al. 2007). Elevated levels are also reported among MCI patients with impending AD as well as asymptomatic patients with FAD-causing mutations (Ringman et al. 2008; Brys and Pirraglia 2007).
20.5.4 Synaptic Biomarkers
Loss of synapses correlates with decrease in neurocognitive function in AD patients (Terry et al. 1991). Biomarkers that reflect this pathology would therefore be desirable. Synaptic proteins such as synaptotagmin, growth-associated protein (GAP-43), synaptosomal-associated protein (SNAP-25), rab3a, and neurogranin have been identified in human CSF, and elevated CSF levels of neurogranin have been reported in both MCI and AD (Davidsson et al. 1999; Thorsell et al. 2010). More research is needed to determine the biomarker potential of synaptic proteins in the CSF.
20.6 Standardization of Biomarkers
The core CSF AD biomarkers have a high diagnostic accuracy for AD, especially in monocenter studies (Andreasen et al. 2001; Tabaraud et al. 2012; Johansson et al. 2011b). However, there is a large variability in measurements between centers and laboratories (Mattsson et al. 2011b). Standardization of pre-analytical and analytical factors is therefore needed before the biomarkers can be implemented in clinical practice. The Alzheimer’s Association has launched a Quality Control Program for CSF biomarkers with the aim of monitoring longitudinal variations and measurements between laboratories (Mattsson et al. 2011b). There are also several other ongoing initiatives, such as the Global Biomarker Standardization Consortium, the CSF-Proteins Working Group of International Federation of Clinical Chemistry and Laboratory Medicine, and the BIOMARKAPD project of the EU Joint Programme in Neurodegenerative Disease Research, that strive towards biomarker standardization (Carrillo et al. 2013). An important step in these standardization efforts is the development of mass spectrometry–based techniques for matrix-independent absolute quantification of Aβ42 in CSF. The first fully validated candidate reference measurement procedure for CSF Aβ42 based on single reaction monitoring (SRM) mass spectrometry has recently been published (Leinenbach et al. 2014). This type of techniques will serve to harmonize different immunoassays with the aim to make uniform cutoff levels across different laboratories.
20.7 CSF Biomarkers in Clinical Research
The CSF biomarkers reviewed above play several important roles in clinical research on AD. They may be used as inclusion criteria in clinical trials of disease-modifying anti-AD drug candidates to verify that the included patients have the pathological changes against which the drug is targeted (Blennow et al. 2010). They may also be used as pharmacodynamic markers or markers of target engagement to test if the drug had the desired biochemical effect in patients on active treatment (Hampel et al. 2014). The markers may also be used in longitudinal studies of healthy volunteers and patients in different stages of AD to learn more about the time course of the different pathogenic processes in AD (Blennow et al. 2010). As an example, proteins secreted into the CSF from activated microglia could be used for determining the role of microglial activation in the AD process. If microglial activation is involved in the disease process and precedes neurodegeneration, a prediction would be that patients on route to AD first drop in CSF Aβ42 (as a sign of Aβ buildup in the brain), then increase in markers of microglial activation, which ultimately is followed by cognitive symptoms and rise in CSF tau (as a sign of neurodegeneration). If microglial activation is a phenomenon downstream of neurodegeneration, such biomarker signals would appear after the rise in CSF tau. Finally, biomarkers for AD-related pathophysiological processes have also proven useful as quantitative traits for genetic analyses (Cruchaga et al. 2013; Andreasson et al. 2014).
20.8 CSF Biomarkers in Other Neurodegenerative Diseases
Pathologies relating to more than one neurodegenerative disorder are not an uncommon finding in postmortem brains. One autopsy study on brains of patients who had a clinical diagnosis of AD found that only 13.6 % of them had pure AD neuropathology, while the others displayed concomitant pathologies such as DLB and TDP (Toledo et al. 2013). This can have an impact in treatment studies targeting a certain pathology, since unknown pathologies may be affecting the outcome. Hence, it is important to find accurate biomarkers relating to pathologies from other diseases than AD.
In the brains of FTD patients, common findings are the neuronal inclusions of tau, TDP-43, or FUS protein (Seelaar et al. 2011). It is a heterogeneous disorder with around one third of the patients having an autosomal dominant variant with mutations in progranulin, microtubule-associated protein tau, or C9orf72. The search for CSF biomarkers for FTD has had limited success. TDP-43 has been measured in the CSF of FTD patients, but appears to originate mainly from blood and be of minor diagnostic importance (Feneberg et al. 2014). Patients with FTD typically have normal or mildly elevated levels of T-tau, but the use of T-tau to distinguish between AD and FTD only gives sensitivity and specificity figures around 80 % (Hampel and Teipel 2004). However, FTD patients have normal levels of P-tau, which can improve the diagnostic separation (Hampel et al. 2004). Further, FTD goes with increased CSF levels of NFL (de Jong et al. 2007).
Patients with DLB and PD belong to a group of diseases called synucleinopathies, based on the findings of α-synuclein in the Lewy bodies that constitute the pathologic hallmark of the diseases. Efforts have been made to quantify α-synuclein in CSF to find a diagnostic biomarker. Lower CSF levels of α-synuclein have been found in patients suffering from α-synucleinopathies (van Dijk et al. 2014; Hong et al. 2010; Mollenhauer et al. 2011; Shi et al. 2011; Hall et al. 2012b; Kasuga et al. 2010), although not in all studies (Spies et al. 2009; Reesink et al. 2010; Noguchi-Shinohara et al. 2009). Altogether, the levels of α-synuclein seem to be reduced in patients with α-synucleinopathies, but with overlap between diagnostic groups. The use of CSF α-synuclein as a biomarker for PD and DLB is also complicated by the finding that α-synuclein seems to reflect neuronal degeneration, with a very marked increase in disorders with marked neuronal loss, such as CJD (Mollenhauer et al. 2008). DLB patients have decreased levels of Aβ42 together with normal or only slightly elevated levels of T-tau in CSF (Parnetti et al. 2008; Zetterberg et al. 2010). Also here, P-tau may be utilized for increased discriminatory power against AD (Parnetti et al. 2008; Vanmechelen et al. 2001).
Due to the heterogenic nature of VaD and also the high degree of uncertainty in the clinical diagnosis of VaD, especially to exclude that a patient fulfilling the clinical criteria for VaD does not have concomitant AD pathology, it is difficult to make definite statements about CSF biomarker changes in patients suffering from this disorder (Roh and Lee 2014). However, findings of an increase in the CSF/serum albumin ratio and an increased CSF NFL level in a patient with a clinical picture suggestive of VaD add support to this diagnosis, while concomitant findings of low CSF Aβ42 and P-tau support a diagnosis of mixed AD/VaD (Blennow et al. 1990; Wallin et al. 1999; Skillback et al. 2013; Sjogren et al. 2001).
As mentioned above, patients with CJD have very high CSF levels of T-tau. As they tend to have normal or slightly elevated CSF levels of P-tau, the ratio of P-tau and T-tau may be used to differentiate CJD from AD (Riemenschneider et al. 2003; Skillback et al. 2014). Indeed, the CJD biomarker profile (T-tau >1,400 ng/L and T-tau/P-tau ratio >25) shows a very high positive likelihood ratio (LR+) for CJD, both in the differential diagnosis against AD (LR + 197) and against other dementias (LR + 109) (Skillback et al. 2014).
20.9 Summary
Three CSF biomarkers reflect the core pathological features of AD: T-tau (neurodegeneration), P-tau (tau hyperphosphorylation and tangle formation), and Aβ42 (plaque pathology). According to revised clinical criteria, these markers may help diagnosing AD also in pre-dementia stages of the disease. At present, their most obvious utility is in clinical trials of novel disease-modifying treatments against AD. In the future, they may help in selecting the right treatment for individual patients by making it possible to assess what molecular pathology that is most likely to cause the patient’s symptoms. Research is currently identifying and validating biomarkers for additional pathophysiological processes, including microglial activation and synapse loss.
References
Agren-Wilsson A, Lekman A, Sjoberg W et al (2007) CSF biomarkers in the evaluation of idiopathic normal pressure hydrocephalus. Acta Neurol Scand 116(5):333–339PubMed
Ahn HJ, Seo SW, Chin J et al (2011) The cortical neuroanatomy of neuropsychological deficits in mild cognitive impairment and Alzheimer’s disease: a surface-based morphometric analysis. Neuropsychologia 49(14):3931–3945PubMed
Albert MS, DeKosky ST, Dickson D et al (2011) The diagnosis of mild cognitive impairment due to Alzheimer’s disease: recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 7(3):270–279PubMedCentralPubMed
Alzheimer A (1987) About a peculiar disease of the cerebral cortex. By Alois Alzheimer, 1907 (Translated by L. Jarvik and H. Greenson). Alzheimer Dis Assoc Disord 1(1):3–8
Andreasen N, Minthon L, Davidsson P et al (2001) Evaluation of CSF-tau and CSF-Abeta42 as diagnostic markers for Alzheimer disease in clinical practice. Arch Neurol 58(3):373–379PubMed
Andreasson U, Lautner R, Schott JM et al (2014) CSF biomarkers for Alzheimer’s pathology and the effect size of APOE varepsilon4. Mol Psychiatry 19(2):148–149PubMedCentralPubMed
Bateman RJ, Xiong C, Benzinger TL et al (2012) Clinical and biomarker changes in dominantly inherited Alzheimer’s disease. N Engl J Med 367(9):795–804PubMedCentralPubMed
Bennett DA, Wilson RS, Schneider JA et al (2002) Natural history of mild cognitive impairment in older persons. Neurology 59(2):198–205PubMed
Bian H, Van Swieten JC, Leight S et al (2008) CSF biomarkers in frontotemporal lobar degeneration with known pathology. Neurology 70(19 Pt 2):1827–1835PubMedCentralPubMed
Blennow K (2004) Cerebrospinal fluid protein biomarkers for Alzheimer’s disease. NeuroRx 1(2):213–225PubMedCentralPubMed
Blennow K, Hampel H (2003) CSF markers for incipient Alzheimer’s disease. Lancet Neurol 2(10):605–613PubMed
Blennow K, Wallin A, Fredman P, Karlsson I, Gottfries CG, Svennerholm L (1990) Blood-brain barrier disturbance in patients with Alzheimer’s disease is related to vascular factors. Acta Neurol Scand 81(4):323–326PubMed
Blennow K, Wallin A, Agren H, Spenger C, Siegfried J, Vanmechelen E (1995) Tau protein in cerebrospinal fluid: a biochemical marker for axonal degeneration in Alzheimer disease? Mol Chem Neuropathol 26(3):231–245PubMed
Related posts:




Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


