Fig. 1
Cross frequency coupling during stimulation with the best frequency (BF) and non-optimal frequencies (nonBF). In each panel the dashed, white oval indicates the CFC frequency ranges used for statistical evaluation (Fig. 2). See main text for further description
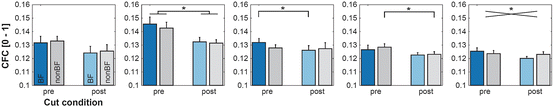
Fig. 2
Statistical evaluation of the influence of dissecting intracortical connections on CFC. The panel order follows Fig. 1. See main text for explanation
3 Results
3.1 Cross-Frequency Coupling Between Measures of Cortical Activity
Stimulation frequencies were split into a BF and a nonBF group as presenting non-optimal frequencies (nonBF) activates horizontal inputs in addition to thalamocortical inputs [5, 8]. For surface LFPs (Fig. 1 left panels) the strongest CFC was observed after 13 Hz fphase and 38 Hz famp (BF stimulation) and at 8 Hz fphase and 33 Hz fAmp (nonBF). The modulation of low and high gamma amplitude by the theta phase appeared very similar to previous reports [3, 13, 16]. Peaks of the CFC comodulograms based on intracolumnar activity (AvgRecCSD) were found at 8 Hz fphase and 33 Hz famp (BF) and at 8 Hz fphase and 28 Hz famp (nonBF) (Fig. 1, column 2). Comodulograms calculated from transcolumnar activity (RelResCSD) peaked at higher frequencies (13 Hz fphase and 38 Hz famp BF and nonBF]; Fig. 1, column 3). Further, the CFC was stronger for BF than for nonBF stimulation (paired t-test: p < 0.032; Fig. 1, column 3). To test how columnar activity influenced transcolumnar processing the fphase was calculated from the AvgRecCSD and famp from the RelResCSD (Fig. 1, column 4). Peaks were found at 8 Hz fphase and 28 Hz famp for both stimulus groups. In contrast, the strongest CFC was found at 13 Hz fphase and 38 Hz famp for BF and nonBF stimulation if fphase was calculated based on RelResCSD and famp based on AvgRecCSD (Fig. 1, column 5). In other words, theta-to-high-beta/gamma coupling was observed for columnar to transcolumnar interactions and beta-to-gamma coupling was detected for transcolumnar to columnar interactions.
3.2 Contribution of Intracortical, Horizontal Connections to Cross Frequency Coupling
To investigate the role of long range processes for CFC, we performed surgical dissections of intracortical, horizontal connections. For the surface LFP theta-to-gamma coupling no significant effects were observed (Fig. 2, panel 1). In contrast, there was a significant reduction of theta-to-low gamma coupling based on the AvgRecCSD (Fig. 2, panel 2, paired t-test: p = 0.041 and p = 0.043 for BF and nonBF). A reduction of RelResCSD based beta-to-gamma coupling was observed after BF stimulation (Fig. 2, panel 3: paired t-test: p = 0.018). Next, we addressed whether cortical cuts modified the coupling of the phase of local activity and the amplitude of global activity (Fig. 2, panel 4). A significant decrease of theta-to-gamma coupling was observed after nonBF stimulation (paired t-test: p = 0.045). The beta-to-gamma coupling from global to local activity influences was also changed with a significant interaction between stimulation frequency and cut condition (Fig. 2, panel 5; RM-ANOVA: p = 0.012).
4 Discussion + Conclusions
Our data indicate that local, columnar cortical activity and global, transcolumnar activity display distinct CFC patterns. While theta-to-gamma coupling was found for local activity an alpha-to-gamma coupling was observed for global activity no matter whether global activity was coordinated in general (RelResCSD to RelResCSD CFC) or global activity tied in local activity (RelResCSD to AvgRecCSD CFC). In line with this idea, a study in human epilepsy patients reported a shift from theta-to-gamma to alpha-to-gamma coupling if larger areas of cortex needed to be coordinated for visual task performance [16].
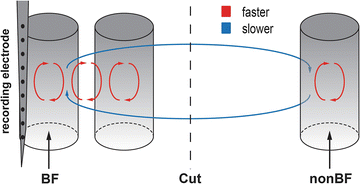
The role of columnar and transcolumnar processes for cross-frequency coupling in cortex can be summarized in a simple model (Fig. 3). Larger neural networks have lower oscillation frequencies than smaller networks [2, 15], thus, local processes, e.g. within a column or between neighboring columns, lead to higher frequency oscillations than global processes, between distant columns. Accordingly, for local activity (AvgRecCSD) presentation of nonBFs leads to a peak in CFC at a lower frequency than stimulation with the BF, as here columnar and transcolumnar processes contribute. For global activity (RelResCSD) no dependency on stimulation frequency was found as long range connections are recruited for all frequencies. Local-to-global interactions were observed at a lower frequency combination than for global-to-local interactions, which might be expected if local activity needs to act on slower, global activity than if global activity acts on fast, local activity. Cortical cuts of long-range connections led to reduced CFC of local activity for all stimuli. This suggests that normal local interactions need global support. CFC of columnar to transcolumnar processes was reduced for nonBFs as long-range horizontal connections which carry information about nonBFs were disrupted. Cortical cuts were more disruptive on BF based RelResCSD CFC than on nonBFs, indicating a directionality of influences such that activity originating from the observed column has a stronger impact on interactions based on horizontal connections than activity originating elsewhere. This might explain the interaction of frequency and cut condition for transcolumnar to columnar influences as BFs depend more on horizontal connections.
 Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
 Dynamic Temporal-Topological Structure of Brain Network Within ADHD
Dynamic Temporal-Topological Structure of Brain Network Within ADHD
 Mind in a Machine: Neural Correlates for Multidimensional Mind Perception
Mind in a Machine: Neural Correlates for Multidimensional Mind Perception
 State Functions Characterizing the Acquisition of New Motor and Cognitive Skills
State Functions Characterizing the Acquisition of New Motor and Cognitive Skills
 Interactions of Two Individual Arm Robots Using Independent Chaos in Recurrent Neural Networks
Interactions of Two Individual Arm Robots Using Independent Chaos in Recurrent Neural Networks
 as Bifurcations Shaped Through Sequential Learning
as Bifurcations Shaped Through Sequential Learning




Related posts:
Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
Dynamic Temporal-Topological Structure of Brain Network Within ADHD
Mind in a Machine: Neural Correlates for Multidimensional Mind Perception
State Functions Characterizing the Acquisition of New Motor and Cognitive Skills
Interactions of Two Individual Arm Robots Using Independent Chaos in Recurrent Neural Networks
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
