Brainstem
The brainstem consists of the medulla oblongata, pons and midbrain. The archaic term ‘bulb’ is applied to the brainstem in compound anatomical names given to nerve fibres originating from, or terminating in, the brainstem (e.g. ‘corticobulbar’ refers to axons that arise in the cerebral cortex and terminate in the brainstem). It is also used clinically to denote the medulla in such terms as ‘bulbar palsy’ and ‘pseudobulbar palsy’, which describe syndromes associated with medullary dysfunction.
The brainstem lies upon the basal portion of the occipital bone (clivus) and is connected to, and largely covered by, the cerebellum. Caudally, the medulla is continuous with the spinal cord just below the foramen magnum. Rostrally, the midbrain is continuous with the diencephalon of the forebrain.
The brainstem contains numerous ascending and descending fibre tracts. Some of these pass throughout its whole length, having their origin in the spinal cord or cerebral hemisphere, respectively; others have their origin or termination within brainstem nuclei. Certain of these brainstem nuclei receive fibres from, or send fibres into, cranial nerves, 10 pairs of which (III–XII) attach to the surface of the brainstem. These are known as the cranial nerve nuclei. In addition, the brainstem contains a complex and heterogeneous matrix of neurones known as the reticular formation, within which a number of individually identified nuclei exist. The reticular formation has several important functions, including control over the level of consciousness, the perception of pain and regulation of the cardiovascular and respiratory systems. It also has extensive connections with the cranial nerve nuclei, with the cerebellum, and with brainstem and spinal motor mechanisms, through which it influences movement, posture and muscle tone. The brainstem contains the cells of origin of monoaminergic (dopamine, noradrenaline, serotonin) neurones that have widespread projections throughout the CNS and are important in sensory, motor, autonomic and cognitive functions.
External features of the brainstem
Dorsal surface of the brainstem
The dorsal surface of the brainstem can be viewed if the overlying cerebellum is removed by cutting the three pairs of nerve fibre bundles, or peduncles, by which it is attached on each side (Figs 9.1, 9.2). On the dorsal surface of the medulla, the midline is marked by a dorsal median sulcus, continuous with that of the spinal cord. In the caudal part of the medulla, the dorsal columns (fasciculi gracilis and cuneatus, containing first-order sensory neurones) continue rostrally from the spinal cord to their termination in the nuclei gracilis and cuneatus, the locations of which are marked by two small elevations, the gracile and cuneate tubercles.
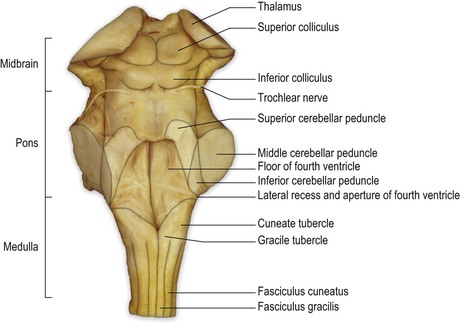
The caudal two-thirds of the medulla contain the rostral continuation of the central canal of the spinal cord and is, therefore, sometimes referred to as the ‘closed’ portion of the medulla. In passing rostrally, the central canal moves progressively more dorsally until, in the rostral medulla, it opens out into the fourth ventricle. This portion is sometimes referred to as the ‘open’ medulla. The floor of the fourth ventricle forms a shallow, rhomboid depression on the dorsal surface of the rostral medulla and the pons. The transition from medulla to pons is not clearly delineated on the dorsal surface of the brainstem but, approximately, the caudal third of the floor of the fourth ventricle constitutes the dorsal aspect of the rostral medulla, while the rostral two-thirds of the ventricular floor is made up of the dorsal aspect of the pons. The fourth ventricle is widest at the level of the pontomedullary junction, where a lateral recess extends to the lateral margin of the brainstem. At this point, a small lateral aperture (foramen of Luschka) provides passage for CSF within the fourth ventricle to reach the subarachnoid space surrounding the brain. The lateral walls of the rostral part of the fourth ventricle are made up of the superior and inferior cerebellar peduncles, connecting the brainstem with the cerebellum. In the rostral pons, the walls converge until, at the pontomesencephalic junction, the fourth ventricle becomes continuous with a small channel, the cerebral aqueduct, which passes throughout the length of the midbrain.
The dorsal aspect of the midbrain is marked by four paired elevations, the superior and inferior colliculi, which are parts of the visual and auditory systems, respectively. The trochlear nerve (IV cranial nerve) emerges immediately caudal to the inferior colliculus.
Ventral surface of the brainstem
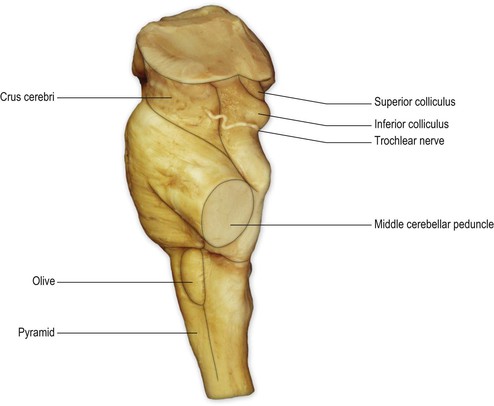
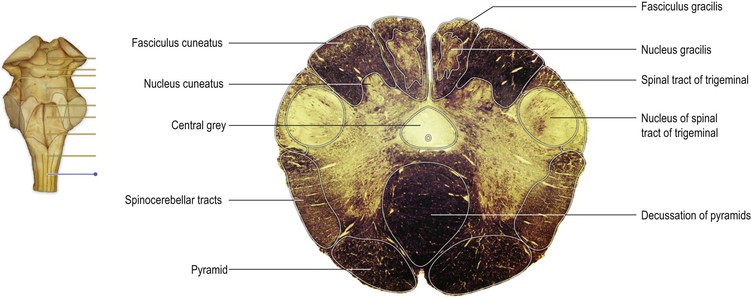
On the ventral surface of the medulla, prominent longitudinal columns, the pyramids, run on either side of the ventral median fissure (Figs 9.3, 9.4). The pyramid gives its name to the underlying pyramidal or corticospinal tract, which consists of descending fibres originating from the ipsilateral cerebral cortex. In the caudal medulla, 75–90% of these fibres cross over in the decussation of the pyramids (Figs 9.3–9.5), partly obscuring the ventral median fissure as they do so, to form the lateral corticospinal tract of the spinal cord. Lateral to the pyramid lies an elongated elevation, the olive, within which lies the inferior olivary nucleus. This has connections primarily with the cerebellum and is involved in the control of movement.
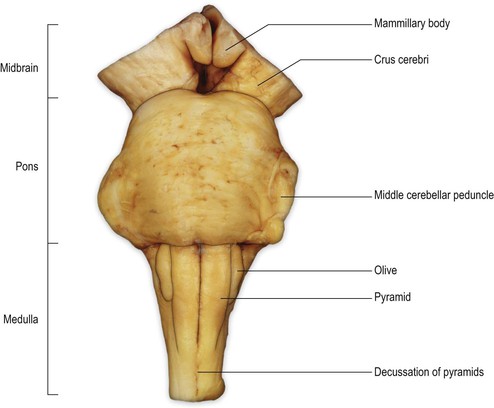
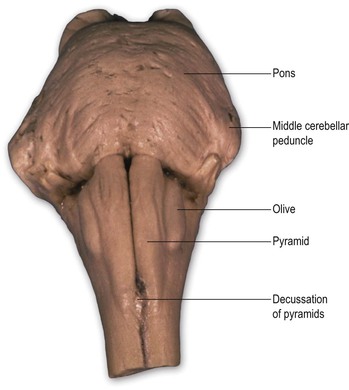
The transition from medulla to pons is clearly delineated on the ventral surface of the brainstem. The ventral part of the pons is dominated by a transverse system of fibres (the transverse pontine fibres or pontocerebellar fibres) that originate from cells in the ventral pons (pontine nuclei) and pass through the contralateral middle cerebellar peduncle to enter the cerebellar hemisphere. The pontine nuclei receive corticopontine fibres from the cerebral cortex (including the motor cortex) and constitute an important connection between cerebral and cerebellar cortices involved in the coordination of movement. The massive system of transverse pontine fibres obscures the underlying corticospinal tract.
The ventral surface of the midbrain consists, on either side, of a large column of descending fibres, the crus cerebri or basis pedunculi. In the midline, the two crura cerebri are separated by a depression called the interpeduncular fossa. The crus cerebri is continuous rostrally with the internal capsule of the cerebral hemisphere (Fig. 1.27) and consists of corticobulbar and corticospinal fibres that have left the cerebral hemisphere via the internal capsule on their way to the brainstem and spinal cord. They are primarily motor in function.
Internal structure of the brainstem
Caudal medulla
At the transition from spinal cord to medulla, the pattern of grey and white matter undergoes considerable rearrangement (Fig. 9.5). The ventral horn becomes much attenuated. The dorsal horn is replaced by the caudal part of the trigeminal sensory nucleus (nucleus of the spinal tract of the trigeminal nerve). The trigeminal sensory nucleus is regarded as the brainstem homologue of the dorsal horn, since it receives primary afferent fibres conveying general sensation from the head, which enter the brainstem in the trigeminal nerve. It is a large nucleus that extends the whole length of the brainstem and into the upper segments of the spinal cord. This latter, caudal part of the trigeminal nucleus is particularly associated with the modalities of pain and temperature. The trigeminal nerve attaches to the pons and, therefore, fibres that terminate in the parts of the trigeminal nucleus caudal to this level descend in a tract (the spinal tract of the trigeminal), which lies immediately superficial to the nucleus.
In the ventral medulla, the majority of fibres of the pyramid undergo decussation and then pass laterally, dorsally and caudally to form the lateral corticospinal tract.
Mid-medulla
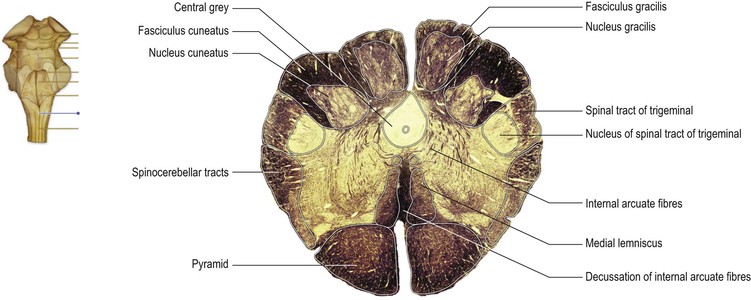
On the ventral surface of the mid-medulla the pyramids are prominent, above their decussation. On the dorsal surface, the ascending fibres of the dorsal columns reach their termination in the gracile and cuneate nuclei, which appear beneath their respective tracts (Fig. 9.6). The dorsal columns consist of first-order sensory neurones; the cell bodies of these neurones lie in the dorsal root ganglia of spinal nerves and have central processes that ascend ipsilaterally through the cord and into the medulla. They terminate in the nuclei gracilis and cuneatus, upon the cell bodies of second-order neurones. The axons of the second-order neurones course ventrally and medially as internal arcuate fibres, decussating in the midline. Thereafter, they turn rostrally forming a distinct tract, the medial lemniscus, which runs through the rostral medulla, the pons and midbrain, to terminate upon third-order neurones in the ventral posterior nucleus of the thalamus.
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


