Cells of the nervous system
The principal functional unit of the nervous system is the nerve cell or neurone. These cells are highly specialised for the encoding, conduction and transmission of information. Neuroglial cells, or glia, are present in the nervous system in even larger numbers than neurones. Glia do not take part directly in information processing but are, nonetheless, crucial for normal neural function. Other cells are also present in the nervous system, such as those forming the walls of blood vessels but, unlike neurones and neuroglia, these are not unique to the nervous system.
The neurone
Neuronal structure
The main structural features, common to all neurones, have been described briefly in Chapter 1. There are, however, numerous variations on the basic plan. The size of the cell body (soma) varies considerably, depending upon its location and function. For example, some interneurones in the CNS have cell bodies as small as 5 µm in diameter, while the cell bodies of motor neurones innervating striated muscle may exceed 100 µm. The size of the cell body is usually correlated with the length of the axon. Therefore, small interneurones usually have short axons, perhaps only a fraction of a millimetre in length. At the other extreme, large motor neurones possess long axons (e.g. those passing from the spinal cord to the muscles of the foot have axons about 1 m in length).
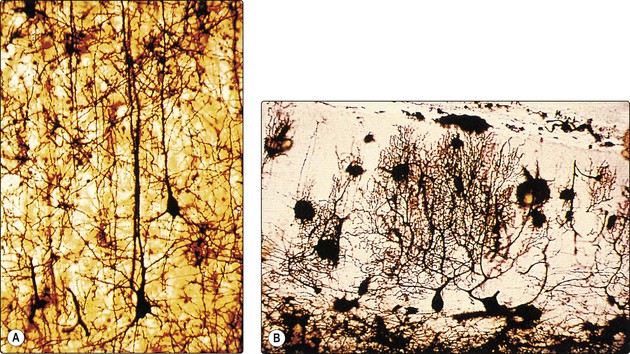
The dendritic arborisation of neurones also shows great variation in the number, size and density of branches, which reflects the organisation of afferent inputs to the cell. For example, pyramidal cells in the cerebral cortex have one or two apical dendrites which course towards the pial surface (Fig. 2.1A), while Purkinje cells in the cerebellar cortex have highly complex, tree-like dendritic arborisations (Fig. 2.1B).
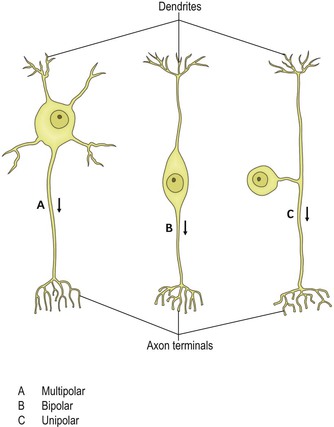
The configuration of the cell body in relation to the dendrites and axon follows one of three basic patterns (Fig. 2.2). Multipolar neurones are by far the most common. Typically, they possess an axon and a number of dendrites that arise directly from the cell body. Bipolar neurones have a centrally placed cell body, from which extend a single dendrite and a single axon. Bipolar neurones occur in the afferent pathways of the visual, auditory and vestibular systems. Unipolar neurones possess a single process emerging from the cell body. This divides into dendritic and axonal branches. Neurones of this type constitute the primary afferents of spinal and some cranial nerves, having their cell bodies in the dorsal root ganglia and sensory ganglia of cranial nerves.
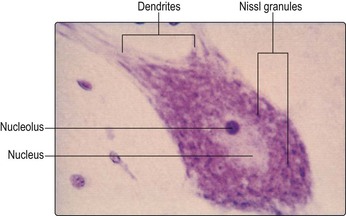
Like most other cells, neurones possess a nucleus. This is usually located in the centre of the cell body and contains the chromosomal DNA. The rest of the intracellular space is occupied by cytoplasm, which contains numerous organelles and inclusions (Fig. 2.3). Many of these are common to cells other than neurones, but some have particular prominence or significance in neurones. Numerous microscopic clumps of Nissl granules (bodies, substance) can usually be seen in nerve cell bodies stained with basophilic dyes. Nissl granules consist of rough endoplasmic reticulum and associated ribosomes. The ribosomes contain RNA (which accounts for the basophilic staining properties) and are the sites of protein synthesis. Nerve cells are highly metabolically active and, therefore, the Nissl granules are often very prominent (Fig. 2.4).
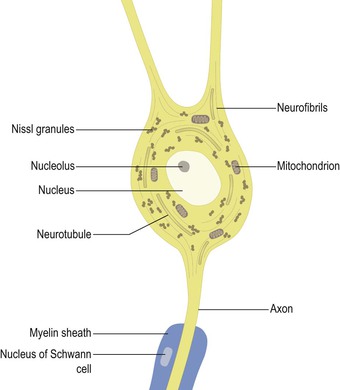
Neurones contain a complex meshwork of structural protein strands called neurofilaments, which are assembled into larger neurofibrils (Fig. 2.3). They also possess a system of neurotubules that are involved in the transport of materials throughout the cell. Transport of materials occurs both away from and towards the cell body (anterograde and retrograde transport, respectively). This phenomenon is exploited in experimental neuroanatomical tracing techniques used to elucidate neuronal connectivity.
Some neurones contain pigment granules. Neuromelanin is a brown-black pigment produced as a by-product of the synthesis of catecholamines. Neuromelanin thus occurs most abundantly in cell groups that utilise catecholamines as their neurotransmitter, notably the pars compacta of the substantia nigra in the midbrain and the locus coeruleus in the pons. Lipofuscin is a yellow-brown pigment that accumulates in some neurones with age.
Each nerve cell is a separate physical entity with a limiting cell membrane. In order for information processing to occur in networks of neurones, therefore, information has to pass between neurones. This occurs at synapses. The basic structure of the synapse has been outlined in Chapter 1. The most common location for synapse formation is between the terminal axonal arborisation of one neurone and the dendrite of another (axodendritic synapse). Other locations are also possible and they give axosomatic, axoaxonal and dendrodendritic synapses. Neurotransmission between neurones occurs by release of specific chemical agents from the presynaptic ending that act upon receptors in the postsynaptic membrane.
Neurotransmitters
Neurotransmitter (transmitter) chemicals are stored in vesicles within the presynaptic ending. A single neurone is thought to release the same transmitter at all its synapses, and all neurones of a particular type, in terms of origin, termination and function, utilise the same transmitter. Numerous transmitter substances have been identified at various sites in the nervous system. It has long been known that, in the periphery, acetylcholine (ACh) is the transmitter between motor neurones and striated muscle. It is also an important transmitter in the autonomic nervous system, being released by preganglionic neurones of both the sympathetic and parasympathetic divisions, and by postganglionic parasympathetic neurones. Furthermore, ACh is a transmitter at numerous, widespread sites within the CNS. Synapses that use ACh as their transmitter are called cholinergic. Some simple amino acids act as neurotransmitters. The most important are glutamic acid (glutamate) and gamma-aminobutyric acid (GABA), which occur widely (forming glutamatergic and GABAergic synapses) and are the principal excitatory and inhibitory transmitters, respectively, in the CNS. Glycine is a transmitter in the spinal cord. Several monoamines are important transmitters. Noradrenaline (norepinephrine) is released by postganglionic sympathetic neurones peripherally as well as at some sites within the CNS. Dopamine and serotonin (5-hydroxytryptamine, 5-HT) also act as transmitters in the brain and spinal cord. Most neurones that utilise monoamines as their neurotransmitter have their cell bodies located in nuclei within the brainstem, e.g. the locus coeruleus (noradrenaline), the substantia nigra (dopamine) and the raphé nuclei (5-HT).
Some small molecule transmitter substances, particularly the amino acids and ACh, are often referred to as fast transmitters because they bring about rapid changes in postsynaptic membrane permeability to certain ions (notably Na+, K+, Ca++ and Cl−) resulting in changes to the postsynaptic membrane potential. Other transmitters have a slower action because they act through the intermediary of a so-called second messenger, such as cyclic adenosine monophosphate (cAMP). Dopamine falls into this category.
A number of peptides (neuropeptides) are stored and released at synapses. These include enkephalin, substance P, cholecystokinin, somatostatin, dynorphin and others. These peptides are often found co-localised in the same neurones as ACh, amino acid or monoamine neurotransmitters and are sometimes called co-transmitters. In many instances, their precise functions are poorly understood. It is believed that some neuropeptides modulate the release, re-uptake and postsynaptic effects of other transmitters and they are, therefore, often referred to as neuromodulators.
The effect of a neurotransmitter must be terminated once it has acted upon the postsynaptic membrane. This is achieved either by enzymic destruction of the transmitter or by its active re-uptake into nerve endings and glia. For example, at the neuromuscular junction (see Fig. 3.6) and at cholinergic synapses, the action of ACh is terminated through hydrolysis by the enzyme acetylcholinesterase (AChE). In contrast, monoamine- and amino acid-mediated transmission are usually terminated by a re-uptake mechanism, although degrading enzymes also exist. Neuropeptides are degraded by peptidases.
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


