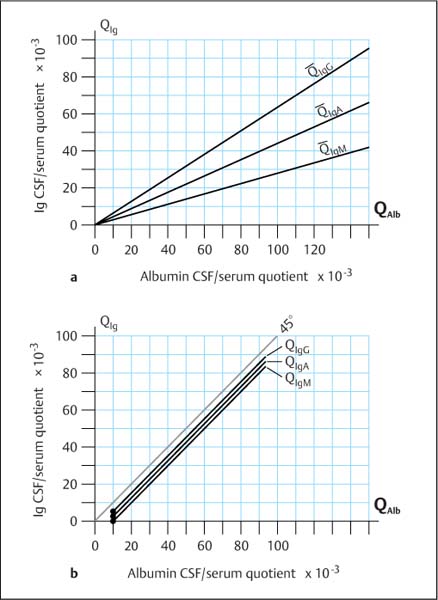
Fig. 3.1 a, b Changes in the CSF concentration of immunoglobulins (QIg) as a function of increasing albumin quotients (QAlb) (blood–CSF barrier dysfunction).
a Mean values of the empirical data for the immunoglobulin CSF/serum quotients with increasing albumin quotients, which may be described as hyperbolic functions (Reiber, 1994 a).
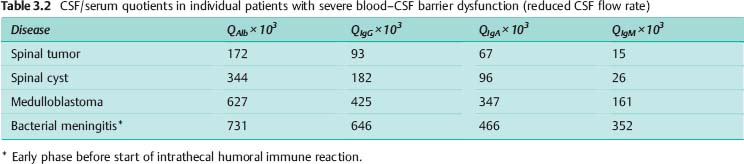
b Experimental simulation of a leakage model in which serum proteins pass in bulk flow from the tissue into the CSF. The immunoglobulin CSF/serum quotients follow a linear function of the increasing albumin quotient. In this in-vitro experiment a patient’s serum was added stepwise to the patient’s CSF sample, and the resulting concentrations of IgG, IgA, IgM, and albumin were measured. Calculation of the CSF/serum quotients for each protein reveals a linear increase, in each case parallel to the 45° line. The empirical data from a large group (a), which show that the molecular-size-dependent discrimination (selectivity) is maintained even in case of the most severe barrier dysfunction, are confirmed by the extreme data from individual patients with different diseases given in Table 3.2. The leakage model (b) cannot explain the reality seen in patients, which is explained quantitatively by the diffusion/flow model.
• Early detection of serum proteins in lumbar CSF: Serum proteins can be detected earlier in lumbar CSF than in ventricular CSF (for references see Reiber, 1994 a). This is explained in the new biophysical model by the steeper local concentration gradient in the spinal subarachnoid space (Reiber, 2003).
• CSF-flow-dependent dynamics of brain proteins: The connection between barrier dysfunction and protein concentration in the CSF is valid for the passage of serum proteins from serum into CSF, but not for the passage of brain proteins into CSF. Whereas the leakage model gives no explanation of the dynamics of brain proteins, the diffusion/flow model provides a sufficient explanation of the dynamics of brain proteins of various origins. Thus, the diffusion/flow model is a more general theory explaining the dynamics of all proteins, both blood- and brain-derived.
The following physiological observations in the context of actual neurological diseases confirm our understanding of the barrier function as changes in the CSF flow rate:
• Leukemia of the CNS: This disease is primarily associated with changes in the trabeculae of the arachnoidea mater. Histopathological studies suggest a reduced CSF flow rate.
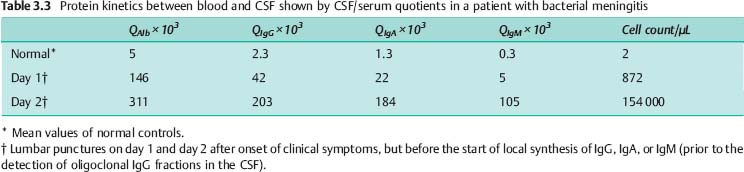
• Purulent bacterial meningitis: This disease is associated with increased CSF viscosity and meningeal adhesions. Post-mortem studies reveal protein complexes and cellular depositions in the arachnoid villi. All these features impede CSF drainage.
• Guillain–Barré syndrome: The high protein concentrations associated with this disease, too, are related to reduced CSF turnover caused by diminished outflow into the veins accompanying the spinal nerve roots, due to swellings in the area around the spinal roots.
• Complete spinal block: In the case of spinal stenosis or complete spinal block, high protein levels are measured in the lumbar CSF caudal to the blockade, despite normal cisternal and ventricular CSF levels. In contrast to bloodderived proteins, proteins originating from the brain, such as transthyretin (formerly called prealbumin), decrease relative to albumin caudal to the blockade. Here, too, the molecular-size-dependent discrimination (selectivity) for protein transfer between blood and CSF is undisturbed.
Brain Proteins in CSF
Site of synthesis of brain proteins. About 20% of the proteins in the CSF originate from the CNS (Thompson, 2005). Of these, only a few are brain-specific (i. e., synthesized exclusively in the CNS). Diagnostically relevant brain proteins come from three main sources (Reiber, 2001; Reiber, 2003):
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


