FIGURE 6.1 Lobes, sulci, and gyri of the lateral aspect of the cerebral hemisphere. The frontal and occipital lobes are finely stippled, the temporal lobe coarsely stippled, and the parietal lobe unstippled.
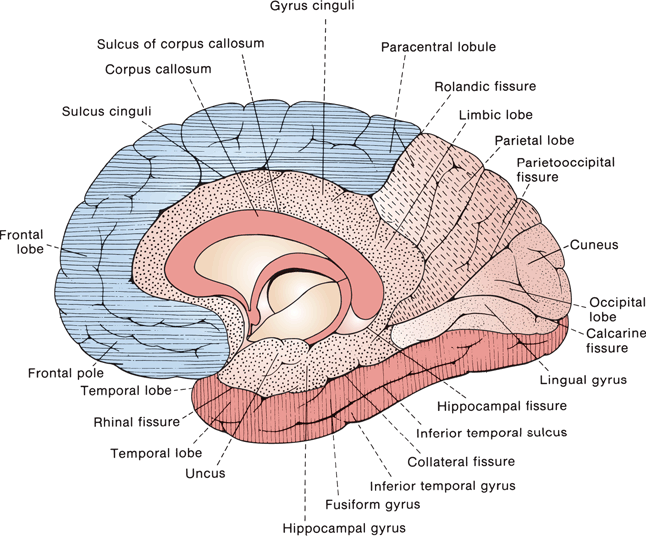
FIGURE 6.2 Lobes, sulci, and gyri of the medial aspect of the cerebral hemisphere. The frontal lobe is lined horizontally and the temporal vertically, the parietal lobe is dashed, the limbic lobe is stippled, and the occipital lobe plain.
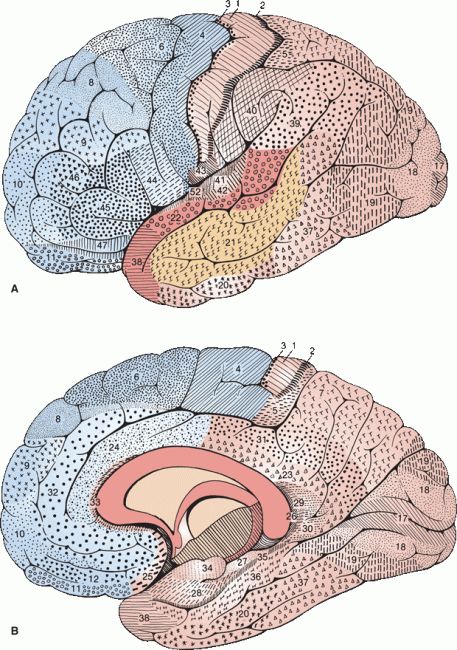
FIGURE 6.3 Areas of the cerebral cortex, each of which possesses a distinctive structure. A. Lateral surface. B. Medial surface. (Modified from Brodmann K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Leipzig: Johann Ambrosius Barth, 1909.)
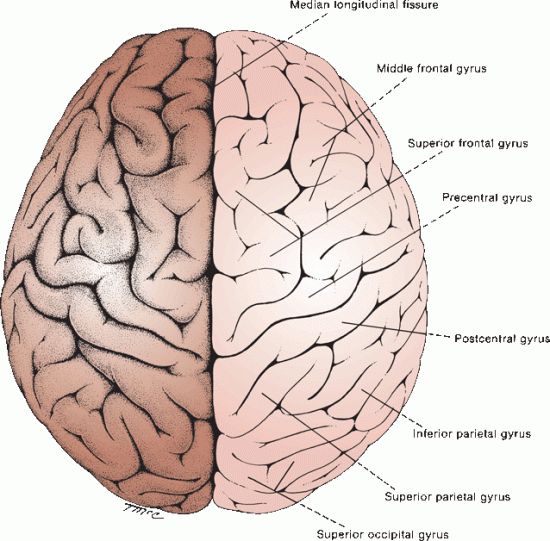
FIGURE 6.4 Gross structure of the cerebral hemispheres as seen from above.
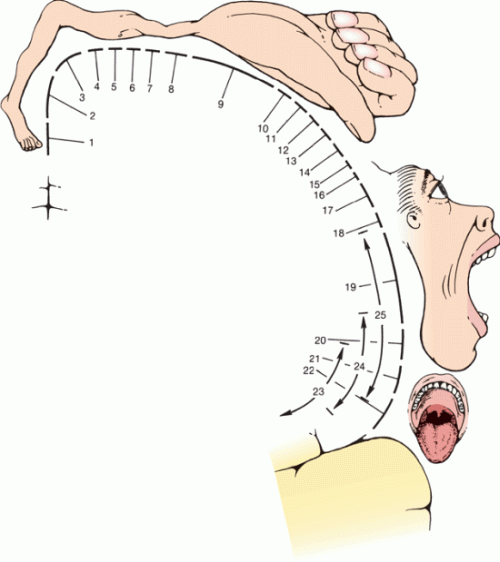
FIGURE 6.5 Motor homunculus, showing the relationship of the motor centers to cortical representation. 1. Toes. 2. Ankle. 3. Knee. 4. Hip. 5. Trunk. 6. Shoulder. 7. Elbow. 8. Wrist. 9. Hand. 10. Little finger. 11. Ring finger. 12. Middle finger. 13. Index finger. 14. Thumb. 15. Neck. 16. Brow. 17. Eyelid and eyeball. 18. Face. 19. Lips. 20. Jaw. 21. Tongue. 22. Swallowing. 23. Mastication. 24. Salivation. 25. Vocalization. (Modified from Penfield W, Rasmussen T. The Cerebral Cortex of Man. New York: Macmillan, 1950.)
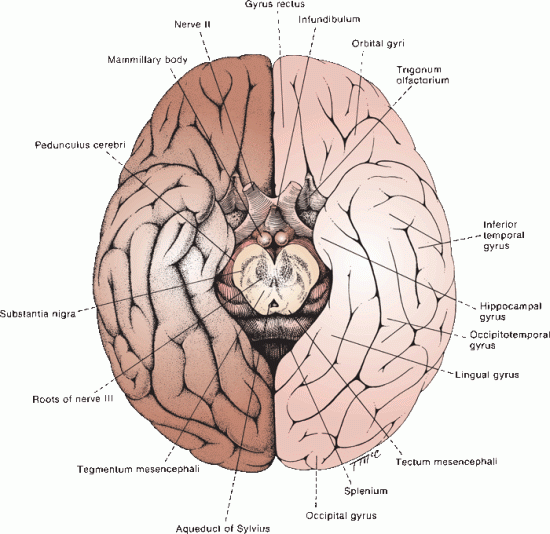
FIGURE 6.6 Base of the human brain.
The parietal lobe lies posterior to the central sulcus, anterior to the occipital lobe and superior to the temporal lobe. An imaginary line drawn between the parieto-occipital sulcus and the preoccipital notch separates the parietal and occipital lobes. An imaginary line extending from the sylvian fissure to the midpoint of the preceding line separates the parietal lobe above from the temporal lobe below. The parietal lobe consists of the following five principal parts: the postcentral gyrus, the superior parietal lobule, the inferior parietal lobule, the precuneus, and the posterior portion of the paracentral lobule. The postcentral gyrus (areas 1, 2, and 3) is the primary sensory cortex; it lies between the central sulcus and the postcentral sulcus. The sensory homunculus depicts the representation of body parts in the primary sensory cortex; it is similar but not identical to the motor homunculus (Figure 6.7). The secondary somatosensory cortex lies in the inferior portion of the postcentral gyrus, abutting the sylvian fissure. The superior parietal lobule is a somatosensory association area that lies posterior to the trunk and upper-extremity segments of the post-central gyrus. The inferior parietal lobule lies posterior to the face and tongue segments of the postcentral gyrus, and it has the following two major components: the supramarginal gyrus, which caps the upturned end of the sylvian fissure; and the angular gyrus, which is at the end of the parallel superior temporal sulcus (Figure 6.1). The inferior parietal lobule is association cortex for somatosensory, visual, and auditory functions. The precuneus is an area of the cortex just anterior to the occipital lobe on the medial hemispheric surface.
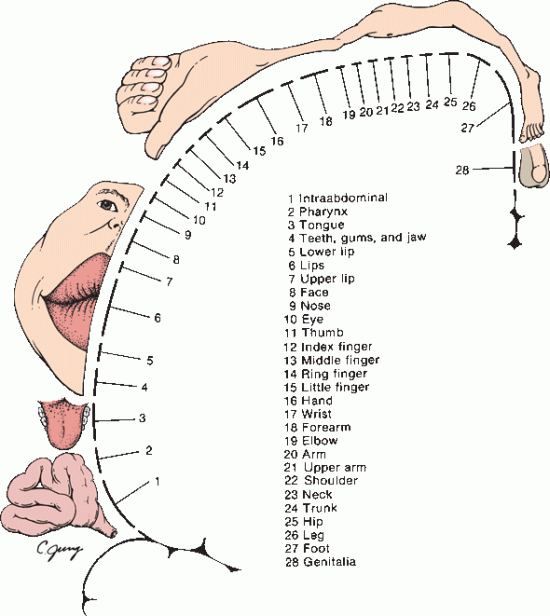
FIGURE 6.7 Homunculus showing cortical sensory representation. (Modified from Penfield W. Rasmussen T. The Cerebral Cortex of Man. New York: Macmillan, 1950.)
The temporal lobe is a tongue-shaped anterior projection that originates as an evagination of the developing cerebral hemisphere; it carries along its central cavity, forming the temporal horn of the lateral ventricle. The temporal lobe lies below the sylvian fissure, extending from the temporal pole to the arbitrary limits of the parietal and occipital lobes. The ventral surface lies on the floor of the middle cranial fossa. The lateral surface has three gyri: the superior, middle, and inferior, which are separated by the superior, middle, and inferior temporal sulci (Figure 6.1). Buried in the sylvian fissure at the posterior end of the superior temporal gyrus on its dorsal surface—running at right angles to the gyrus and stretching toward the medial geniculate body—are the transverse temporal gyri (of Heschl). The transverse temporal gyri are the primary auditory cortex (areas 41 and 42). Immediately adjacent to the primary auditory cortex is the auditory association cortex (area 22); in the dominant hemisphere part of this is the Wernicke’s speech area. The planum temporale lies just behind the Heschl gyri and is part of Wernicke’s area. The planum temporale is larger in the left hemisphere in most individuals and is probably related to cerebral dominance for language. On the base of the temporal lobe, the inferior temporal gyrus is continuous medially with the lateral occipitotemporal gyrus. The occipitotemporal sulcus separates the lateral occipitotemporal (inferior temporal) gyrus from the medial occipitotemporal (fusiform) gyrus. Medial to the fusiform gyrus, separated by the collateral sulcus, is the parahippocampal (hippocampal) gyrus, part of the limbic lobe. Posterior to the isthmus of the cingulate, the parahippocampal gyrus stretches toward the occipital pole and becomes the lingual gyrus.
The occipital lobe is only a small part of the dorsolateral surface of the hemisphere, but it occupies a large triangular field on the medial aspect of the brain between the parietal and temporal lobes. The calcarine fissure separates the medial surface of the occipital lobe into the cuneus above and the lingual (medial occipitotemporal) gyrus below. The occipital lobe is the visual cortex (areas 17, 18, and 19). The cuneus forms the upper bank, and the lingual gyrus the lower bank, of the calcarine cortex.
The limbic lobe is sometimes considered a separate lobe of the brain, more because of its function than its anatomy. Components of the limbic lobe include the following: the hippocampus, which lies deep in the medial temporal lobe and becomes continuous with the fornix; the mamillary bodies (part of the hypothalamus); the anterior nucleus of the thalamus; the cingulate gyrus; and the parahippocampal gyrus. As with several other central nervous system (CNS) structures, the limbic lobe morphologically is a C-shaped structure. It begins anteriorly and superiorly in the paraterminal gyrus and subcallosal area beneath the rostrum of the corpus callosum. The body of the C is formed by the cingulate gyrus, which merges at the isthmus of the cingulate into the parahippocampal (hippocampal) gyrus. The end of the C is the hippocampal formation. The cingulate gyrus lies just above the corpus callosum. The parahippocampal gyrus begins at the isthmus of the cingulate and runs to the temporal tip, lying between the collateral sulcus and the hippocampus; it curls around the hippocampal fissure to form the uncus. The hippocampal formation is composed of the hippocampus proper (Ammon’s horn), the dentate gyrus, and the subiculum. When not regarded as part of the limbic lobe, the anterior and posterior parts of the cingulate gyrus are considered parts of the frontal and parietal lobes, respectively. The parahippocampal gyrus and hippocampal formation are considered part of the temporal lobe. The structures of the limbic lobe are connected in Papez circuit (cingulate gyrus → parahippocampal gyrus → hippocampus → fornix → mamillary body → anterior nucleus of the thalamus → cingulate gyrus).
The rhinencephalon (nose brain) is a primitive, basal forebrain region involved with olfaction and emotion that is closely related to the limbic lobe. It consists of the olfactory bulbs and tracts, the olfactory stria, olfactory trigone (olfactory tubercle, anterior perforated substance and diagonal band of Broca), the piriform lobe (uncus, entorhinal area and limen insulae), and part of the amygdala. The hippocampal formation is sometimes included as part of the rhinencephalon.
CORTICAL LAYERS
The cerebral cortex begins as an outpouching of the rostral end of the neural tube, and it culminates as a complex cellular layer that covers the surface of the brain. After formation of the marginal and mantle layers, cells migrate from the marginal layer to form the cerebral cortex. Migrational defects are a common cause of congenital brain malformations, such as gray matter heterotopias. Between the sixth and eighth month of fetal life, the migrating cells reach the cortex and become organized into strata, which eventually become the cortical layers. The cortex covers the gyri and convolutions and dips into the fissures and sulci. About one-third is on the exposed surface, and the rest is buried in the fissures and sulci. There are about 15 to 30 billion nerve cells in the cortex. Its thickness varies from 4.5 mm in the precentral gyrus to 1.3 mm near the occipital pole.
Most of the cortical mantle has six identifiable layers; some areas of the brain have less (Figure 6.8). Six-layered cortex is referred to as neocortex, isocortex, or heterogenetic cortex. The six layers, from superficial to deep, are as follows: (1) molecular (plexiform), (2) external granular, (3) external pyramidal, (4) internal granular, (5) internal pyramidal (ganglion), and (6) multiform. The molecular layer is most superficial, covered by pia. It consists of a dense tangle of fibers composed of dendrites of deeper lying cells. Pyramidal cells are sparse and small. Layer 2, the external granular layer, is made up of small, densely packed neurons. Layer 3, the external pyramidal layer, consists of medium to large pyramidal-shaped neurons. It is sometimes subdivided into a superficial layer of medium pyramidal cells and a deep layer of large pyramidal cells. Layer 4, the internal granular layer, consists of many small, multipolar granule cells with short axons and scattered small pyramidal cells. Granule cells are most numerous in this layer. Layer 5, the internal pyramidal (ganglion cell) layer, consists of medium and large pyramidal cells, among which are the largest neurons found in the cortex. In the precentral gyrus, this layer contains the giant pyramidal cells of Betz, the neurons whose axons form the corticospinal and corticobulbar tracts. The deepest cortical layer is the multiform layer, which consists of polymorphic cells whose short axons enter the subjacent white matter.
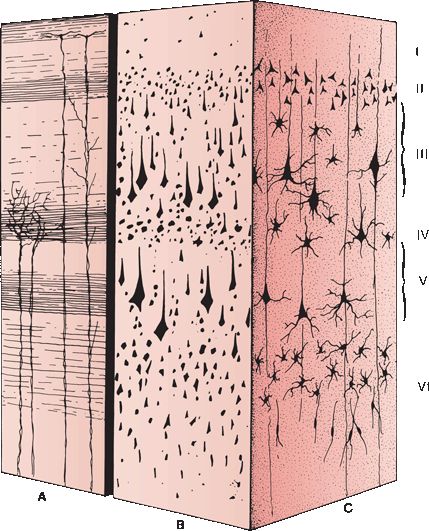
FIGURE 6.8 Cell layers and fiber arrangement of the cerebral cortex. A. Weigert stain. B. Nissl stain. C. Golgi stain. Layers: I. Molecular layer II. External granular layer III. External pyramidal layer IV. Internal granular layer V. Internal pyramidal layer VI. Multiforme layer.
Isocortex is found in the neopallium, which makes up about 90% of the cortical surface. Severe compromise of brain energy supplies, such as in hypoxia, ischemia, or hypoglycemia may lead to selective destruction of certain cortical layers, mainly the third—a condition termed cortical laminar necrosis. The archipallium and paleopallium both have three-layered cortex, referred to as allocortex.
Different areas of the cortex have characteristic appearances, with differences in the overall thickness of the cortical layer, the thickness and arrangement of specific cellular layers, the cell structure, the number of afferent and efferent myelinated fibers, and the number and position of white stria. How regional differences in the cytoarchitecture correlate with differences in function remains a matter of conjecture. Maps based on differences in cellular structure are referred to as cytoarchitectonic and on differences in fiber structure as myelotectonic. The best known cytoarchitectonic map is that of Brodmann (Figure 6.3). Modern imaging and the use of other cortical markers may lead to a newer generation of more accurate maps.
The cortex sends and receives fibers to and from other areas of the brain. Layer 4 contains a dense horizontal band of fibers—the external band of Baillarger. This band contains the terminal ramifications of the thalamocortical projections from the specific thalamic relay nuclei. The external band of Baillarger is particularly prominent in the calcarine cortex, forming a grossly visible white stripe—the line or band of Gennari—that gives the striate cortex its name. The specific thalamic sensory nuclei synapse in layer 4. The external band of Baillarger is made up of the terminal ramifications of thalamic nuclei that subserve specific sensory modalities, such as vision and exteroceptive sensation. In contrast, the nonspecific thalamic nuclei (reticular, intralaminar) project diffusely to all layers of the cortex.
Isocortex can also be simply divided into supra-granular and infragranular layers. Layers above layer 4 (the dense internal granular layer) are supra-granular; those below layer 4 are infragranular. The supragranular cortex (primarily layer 2 and layer 3) is highly differentiated and phylogenetically recent. Supragranular afferents and efferents are primarily associative; they are concerned with higher-level integrative functions and corticocortical connections. The infragranular cortex is more primitive. It is well developed in lower forms, and it primarily sends descending projection fibers to lower centers. Six-layered isocortex is formed essentially by the presence of supragranular cortex atop three-layered allocortex. The supragranular layers are not present in the archipallium and paleopallium.
Isocortex may be either homotypical, in which six layers can be easily discerned, or heterotypical, in which lamination is less obvious. The cortex can also be divided into granular and agranular types. In agranular cortex, the granule cell layers are poorly developed, while the pyramidal cell layers are prominent. Agranular cortex is characteristic of the precentral gyrus. Granular cortex (koniocortex) is thin and contains dense granule cell layers; the pyramidal cell layers are less conspicuous. Granular cortex is characteristic of areas that receive heavy afferent input, such as the calcarine cortex. There is a striking paucity of granule cells in the agranular cortex, for example, the motor strip, and a paucity of pyramidal cells in the granular cortex, for example, the primary sensory areas. Koniocortex is seen only in areas that receive projections from the specific thalamic relay nuclei. Cortical areas that receive thalamocortical projections from the specific thalamic relay nuclei therefore have the following two morphologic characteristics: granular type cortex and a prominent external band of Baillarger.
In addition to its horizontal, laminated organization, the cortex is also organized vertically into columns. Neurons subserving the same modality and with similar receptive fields are organized into vertical rows that extend from the cortical surface to the white matter, which is referred to as cortical columns. The vertical column organization is particularly prominent in the occipital, parietal, and temporal lobes.
Beneath the cortical mantle of gray matter lies the white matter, which consists of association, commissural, and projection axons—as well as glial cells and blood vessels. The association and commissural fibers connect one area of the cortex with another. Association fibers connect cortical areas within the same hemisphere; commissural fibers connect to areas in the opposite hemisphere. Association and commissural fibers come primarily from the supragranular cortex (layer 1 to layer 3). Projection fibers connect the cortex with lower centers (Figures 6.9 and 6.10). Projection fibers arise primarily from the infragranular cortex (layer 5 and layer 6) and go to lower centers of the nervous system. The corticospinal tract is composed of projection fibers that arise from neurons in the deeper layers of the precentral gyrus. The number of projection fibers is surprisingly small in comparison to the total number of neurons in the cortex.
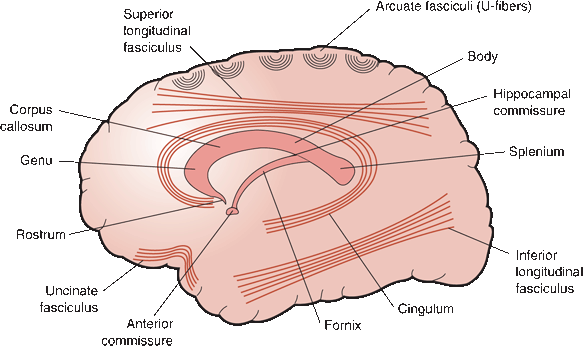
FIGURE 6.9 Sagittal view demonstrating short association fibers (arcuate or U-fibers), long association bundles, and major commissures.




