CHAPTER 55 HEADACHE PATHOGENESIS
Headache is a broad term that encompasses most of the pain syndromes that involve the head and upper neck. Headache is much more than migraine alone, and facial pain is often included in the discussion of headache. The common feature of all these pain syndromes is that they involve the regions of the body innervated by the trigeminal nerve and the upper cervical nerves, as opposed to nonheadache pain syndromes, which involve the spinal nerves. This chapter discusses the pathogenesis of headache. We discuss the tissues in the periphery and the regions of the brain that are involved in generating and maintaining headache pain, with an emphasis on the pathophysiology of migraine. This emphasis on migraine is due to the advances that have been made in animal models of headache and functional imaging studies of patients with migraine.
Migraine is a primary headache disorder arising from a brain dysfunction that leads to activation of the trigeminal vascular system. This review discusses developments in the understanding of the cellular and molecular aspects of migraine. Migraine is more than a pain disorder—there are multiple stages of a migraine attack. The prodrome may begin as early as 3 days before an attack; in some patients (≈14%) this leads to a visual or sensory aura. The prodrome phase is followed by the pain phase, which, untreated, can last from 4 to 72 hours. This is followed by the resolution phase, which is characterized by alleviation of the pain and a return to normal activity. Even during this interictal phase, auditory habituation1 and threshold measurements of magnetic stimulation of the cortex2 are different in migraineurs compared with healthy subjects; the blink reflex, however, is not different.3
ANATOMY OF HEADACHE
Most animal research into migraine has focused on activation of dural nociceptors using electrical current or proinflammatory chemicals. Dural blood vessels and the dura itself are innervated by sensory neurons whose cell bodies are in the trigeminal ganglion. A single neurite emerges from the cell bodies of the sensory neurons in the trigeminal ganglion. This neurite bifurcates and projects out to the periphery and into the dorsal horn of the midbrain. Because sensory neurons in the trigeminal ganglion have only one neurite, both projections are classified as axons. Normally, we associate the direction of information transmission along this neuron from the periphery to the central nervous system, but glutamate and neuropeptides can also be released in the periphery. This can contribute to sensory trigeminal activation, blood vessel vasodilation, and plasma protein extravasation.4 Glutamate and neuropeptides, such as calcitonin gene–related peptide, can also be released in the trigeminal ganglion itself.5–7 These sites of release are important for the strong sensory activation required for the initiation of the migraine attack and as potential targets of migraine headache treatments. Significant in headache, the large blood vessels of the meninges are mainly innervated by Aδ and C type sensory fibers, which contain glutamate as well as neuropeptides.
The fifth cranial nerve arises from the trigeminal ganglion and transmits all the somatic sensory information from the head, face, and dura to the trigeminal nucleus caudalis in the central nervous system. The trigeminal nerve has three divisions: ophthalmic, mandibular, and maxillary. Anterior structures of the head and face are innervated by the ophthalmic (first) division. Posterior regions of the head and neck are innervated by the upper cervical nerves. The trigeminal nerve enters the brainstem at the pontine level of the hindbrain and terminates in the trigeminal nucleus of the brainstem. Nociceptive input from the cervical nerves also activates the neurons in the trigeminal nucleus caudalis, which extends to the C2 spinal segment.8
The trigeminal nucleus is composed of the principal trigeminal nuclei and spinal trigeminal nuclei (subdivided into three regions: the nucleus oralis, the interpolaris, and the nucleus caudalis).9 The dorsal horn of the brainstem spinal trigeminal nucleus has a laminar structure homologous to the dorsal horn of the spinal cord. The primary afferents terminate in laminae I and II and, to a lesser extent, the deeper laminae III to V. The neurons of the deeper lamina project to the thalamus in the trigeminothalamic tract, which is homologous to the spinothalamic tract. Second-order neurons from the trigeminal spinal nuclei form the trigeminothalamic tract and project to other midbrain structures, such as the periaquaductal gray, as well as to the thalamus and hypothalamus (Fig. 55-1).
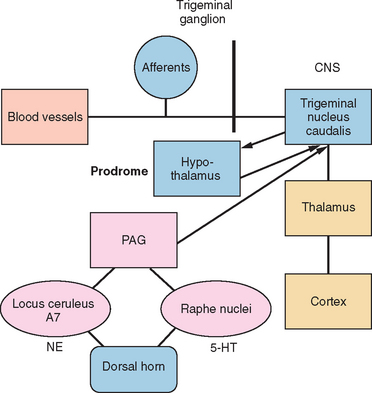
HEADACHE PAIN
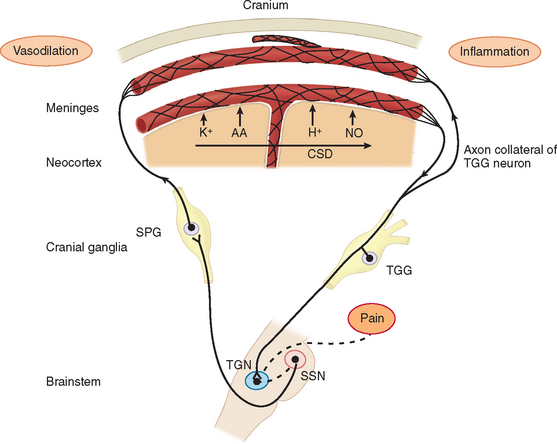
Migraine pain is caused by trigeminal neurovascular activation.10 Neural events result in activation of nociceptive afferents and dilation of blood vessels on the dura. In patients with migraine accompanied by aura, cortical spreading depression is the neural event that activated the nociceptors on the dura. Cortical spreading depression is really a spreading wave of cortical spreading excitation followed by long-lasting depression. In migraine patients without aura, the mechanism for the activation of the dural afferents is unknown.
Studies in humans and research on headache models in rats have elucidated three main components involved in headache pain: (1) the cranial blood vessels, (2) the trigeminal innervation of the vessels, and (3) the reflex connections of the trigeminal system with the cranial parasympathetic system.11 The key pathway for headache pain is trigeminovascular input from the meningeal vessels and the meninges.12 Brain imaging studies suggest that important modulation of the trigeminovascular nociceptive input stems from the dorsal raphe nucleus, the locus ceruleus, and the nucleus raphe magnus.13,14
PERIPHERAL MECHANISMS AND NOCICEPTOR ACTIVATION
Before the contribution of neurogenic inflammation during the pain phase of migraine was demonstrated, the predominant theory of migraine was Harold Wolff’s vascular theory. Wolff postulated that migraine was a vascular disorder caused by a tightening (constriction) and sudden opening (dilation) of the blood vessels in the head. Research has shown that changes in blood flow are due to activation of afferents and the brain. This led to the neurogenic theory of migraine. During neurogenic inflammation, trigeminal afferent activation is accompanied by the release of vasoactive neuropeptides, including calcitonin gene–related peptide, substance P, and neurokinin A from the nerve terminals.10,15,16 These mediators produce vasodilation, sensitization of the nerve terminals, and extravasation of fluid into the perivascular space around the dural blood vessels (plasma protein extravasation) (Fig. 55-2). The nerve terminals become further excited due to a positive feedback loop. The afferents excite the blood vessels and other trigeminal nerve endings through glutamate and peptide release, and then the blood vessels further excite the sensitized afferent terminals. These afferents project in the central nervous system and terminate in the upper lamina of the dorsal horn. Some project to the deeper lamina.17,18 This strong sensory stimulation causes the induction of c-fos (an immediate-early gene product) in the secondary sensory neurons of the trigeminal nucleus caudalis in the brainstem.19 Substance P and calcitonin gene–related peptide further amplify the trigeminal terminal sensitivity by stimulating the release of bradykinin and other inflammatory mediators from mast cells. Prostaglandins and nitric oxide (a diffusible gas that acts as a neurotransmitter) are both endogenous neuromodulators that can sensitize nociceptors and the secondary sensory neurons in the trigeminal nucleus caudalis. There is evidence that inflammatory mediators, such as prostaglandin release in the central nervous system by microglia, may be more important than its release on the dura.20 Microglia are important modulators on sustained pain in chronic pain conditions.21
< div class='tao-gold-member'>
Related posts:
 AUTISM AND ATTENTION DEFICIT/HYPERACTIVITY DISORDER
AUTISM AND ATTENTION DEFICIT/HYPERACTIVITY DISORDER
 PRINCIPLES OF NEUROPSYCHOMETRIC ASSESSMENT
PRINCIPLES OF NEUROPSYCHOMETRIC ASSESSMENT
 TRIGEMINAL AUTONOMIC CEPHALALGIAS: CLUSTER HEADACHE AND RELATED CONDITIONS
TRIGEMINAL AUTONOMIC CEPHALALGIAS: CLUSTER HEADACHE AND RELATED CONDITIONS
 ANATOMY AND PHYSIOLOGY OF CEREBRAL AND SPINAL CORD CIRCULATION
ANATOMY AND PHYSIOLOGY OF CEREBRAL AND SPINAL CORD CIRCULATION
 TUMORS OF THE SPINAL CORD
TUMORS OF THE SPINAL CORD
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


