Fig. 1
Stimulation pattern
3 Result
3.1 STDP Was Enhanced by the Application of Eserine
First, in order to investigate the influence of endogenous ACh on the induction of STDP, pairing stimulation was applied in the presence of different concentrations (2 and 10 μM) of eserine. When an STDP-inducing stimulus in pre-post timing was applied, LTD in the control condition (70.1 ± 7.1 %, n = 5, P < 0.05) was changed to LTPs in the presence of 2 μM eserine (111.9 ± 2.1 %, n = 5, P < 0.01, P < 0.01 vs. control) and 10 μM eserine (120.5 ± 4.8 %, n = 6, P < 0.01, P < 0.01 vs. control) respectively as shown in Fig. 2. These results show that STDP was shifted toward potentiation by the activation of AChRs. Next, in order to clarify the effect of GABAA receptor (GABAAR) activation on cholinergically induced STDP, picrotoxin (25 μM) was added to ACSF in the presence of eserine 2 μM. When the STDP-inducing protocol in pre-post timing was applied only in the presence of 2 μM eserine, LTP was observed as in Fig. 2, and the LTP was significantly enhanced in the additional application of picrotoxin (179.3 ± 9.1 %, n = 5, P < 0.01, P < 0.01 vs. eserine 2 μM only; Fig. 3a). As these results indicate that GABAAR contributed to the cholinergically-induced STDP, next the dependence of eserine on IPSCs were confirmed. The magnitude of IPSCs in control condition was significantly enhanced in the presence of 2 μM eserine (1.28 ± 0.06 %, n = 4, P < 0.01, P < 0.01 vs. control; Fig. 3b, c). These results show that interneurons were strongly activated by the application of eserine, therefore the cholinergically induced STDP was regulated by interneurons.
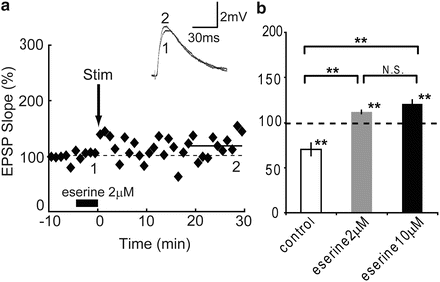
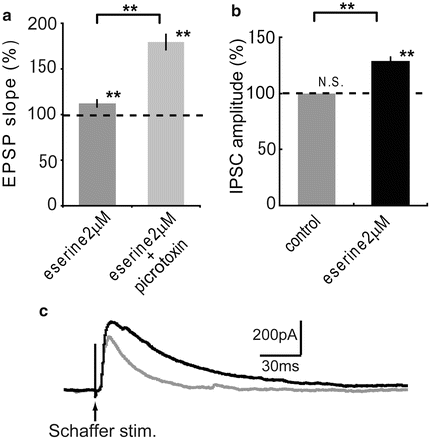
Fig. 2
Eserine effect on STDP
Fig. 3
Eserine effect on IPSC
3.2 Cholinergically-Induced STDP Was Contributed by mAChR and nAChR
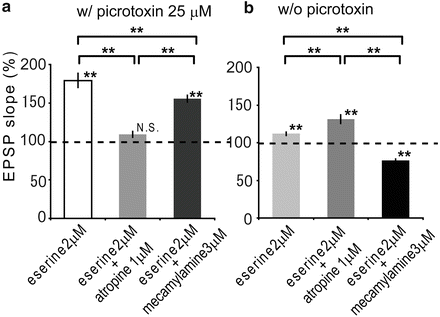
As STDP was enhanced by the application of eserine, in second, the dependence of each AChR was clarified. When an STDP-inducing protocol in pre-post timing was applied in the presence of eserine 2 μM and picrotoxin 25 μM, LTP was observed as in Fig. 4a. When atropine at 1 μM was added to the eserine and picrotoxin treated ACSF, LTP was abolished (109.2 ± 4.0 %, n = 6, N.S., P < 0.01 vs. eserine 2 μM only; Fig. 4a), while LTP was significantly reduced in the presence of 3 μM mecamylamine instead of atropine (155.6 ± 4.3 %, n = 5, P < 0.01, P < 0.05 vs. eserine 2 μM only, P < 0.01 vs. eserine 2 μM + atropine 1 μM). According to the results, cholinergically-induced STDP was regulated by the activation of mAChRs more effectively than nAChRs on pyramidal neuron. Furthermore, in order to investigate the effect of AChRs on interneurons, similar experiments were performed in the absence of picrotoxin. LTP induced in the presence of eserine 2 μM (Fig. 2) was significantly enhanced with additional application of atropine 1 μM (131.4 ± 5.7 %, n = 5, P < 0.01, P < 0.01 vs. eserine 2 μM only; Fig. 4b), while STDP was changed to LTD in the presence of 3 μM mecamylamine instead of atropine (76.5 ± 2.0 %, n = 5, P < 0.01, P < 0.01 vs. eserine 2 μM only, P < 0.01 vs. eserine 2 μM + atropine 1 μM). Therefore cholinergically-induced STDP was regulated by AChRs on interneurons. Taken together, these findings show that the activation of mAChRs contributed more effectively than nAChRs not only on pyramidal neurons but also on interneurons.
 Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
 Dynamic Temporal-Topological Structure of Brain Network Within ADHD
Dynamic Temporal-Topological Structure of Brain Network Within ADHD
 Mind in a Machine: Neural Correlates for Multidimensional Mind Perception
Mind in a Machine: Neural Correlates for Multidimensional Mind Perception
 State Functions Characterizing the Acquisition of New Motor and Cognitive Skills
State Functions Characterizing the Acquisition of New Motor and Cognitive Skills
 Interactions of Two Individual Arm Robots Using Independent Chaos in Recurrent Neural Networks
Interactions of Two Individual Arm Robots Using Independent Chaos in Recurrent Neural Networks
 as Bifurcations Shaped Through Sequential Learning
as Bifurcations Shaped Through Sequential Learning




Related posts:
Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
Dynamic Temporal-Topological Structure of Brain Network Within ADHD
Mind in a Machine: Neural Correlates for Multidimensional Mind Perception
State Functions Characterizing the Acquisition of New Motor and Cognitive Skills
Interactions of Two Individual Arm Robots Using Independent Chaos in Recurrent Neural Networks
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
