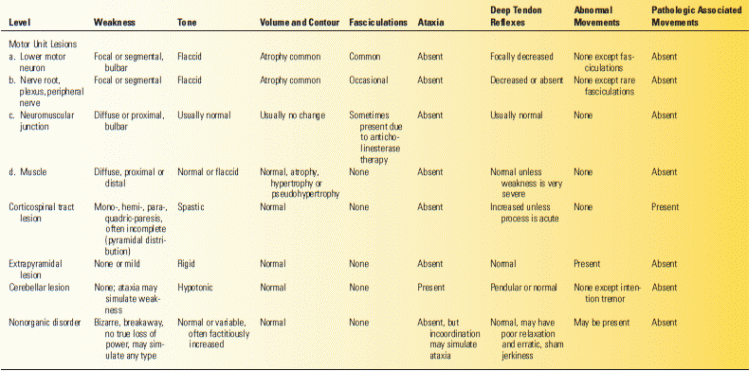
FIGURE 22.1 The most important descending pathways that act upon the anterior horn cell of the spinal cord (final common pathway).
These levels are not individual motor systems and do not normally act individually or separately. Anatomists continue to have difficulty in even defining the constituents of some of these levels (e.g., the corticospinal or pyramidal vs. the basal ganglion or extrapyramidal). These levels are components of the motor system as a whole; each is part of the complex motor apparatus. Each contributes its share to control of the lower motor neuron on which, as the final common pathway, all motor control systems converge. Disease at each of these levels causes characteristic signs and symptoms (Table 22.1). Some disorders of the motor system may involve more than one level. In addition, all purposeful movements are guided by a constant stream of afferent impulses that impinge on various levels of the motor system. Sensory and motor functions are interdependent in the performance of volitional movement, and it is not possible to consider the motor system apart from the sensory system. Impairment of sensation may affect all aspects of motion—volitional, reflex, postural, tonic, and phasic.
TABLE 22.1 Changes in Motor Function
A brief sketch of the organization of the motor system may help to lay a foundation before discussing each level in detail (Figures 22.1 and 22.2). The pre-motor and supplementary cortices control the planning and preliminary preparation for movements, which the primary motor cortex in the precentral gyrus then executes. The primary motor cortex also receives input from the basal ganglia and the cerebellum (Figure 22.2). The corticospinal (pyramidal) and corticobulbar tracts arise from the precentral gyrus, descend through the corona radiata, and enter the posterior limb of the internal capsule. The internal capsules merge in their descent with the cerebral peduncles, which form the base of the midbrain. Corticobulbar fibers terminate in the lower brainstem on cranial nerve nuclei and other structures. Corticospinal fibers aggregate into compact bundles, the pyramids, in the medulla. At the level of the caudal medulla, 90% of the pyramidal fibers decussate to the opposite side and descend throughout the spinal cord as the lateral corticospinal tract. About 10% of the corticospinal fibers descend ipsilaterally in the anterior corticospinal tract and decussate at the level of the local spinal synapse. Pyramidal fibers preferentially innervate certain lower motor neuron groups.
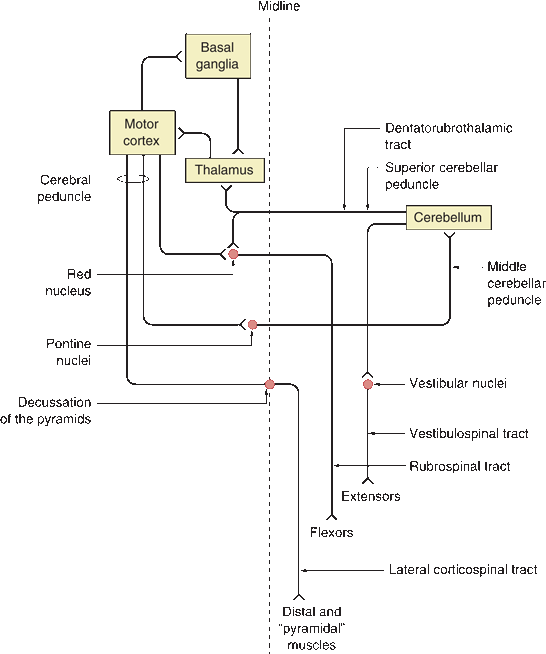
FIGURE 22.2 Major connections of the motor system. Note feedback loops between cortex and cerebellum and cortex and basal ganglia. (Modified from Campbell WW, Pridgeon RP. Practical Primer of Clinical Neurology. Philadelphia: Lippincott Williams & Wilkins, 2002.)
Descending motor system fibers send collaterals to other structures to help control and coordinate movement. These structures in turn project back to the cortex to form feedback loops that ensure coordinated interactions between the suprasegmental motor systems (Figure 22.2). The thalamus, especially the ventral lateral (VL) and the ventral anterior (VA) nuclei, serves as the relay station for projections from the other centers back to the cortex. The VL projects predominantly to the primary motor cortex and the VA to the premotor regions.
As fibers from the motor cortex run downward through the internal capsule, they send collaterals to the basal ganglia. Fibers from the basal ganglia project to VA and VL, which in turn project to the cortex, creating a feedback loop. The substantia nigra also projects to the striatum and influences its activity. The motor cortex and cerebellum are also part of a circuit. The pontine nuclei lie scattered among the descending motor and crossing pontocerebellar fibers in the basis pontis. Corticopontine fibers synapse on pontine nuclei, which then give rise to pontocerebellar fibers that project across the midline to the contralateral cerebellar hemisphere through the middle cerebellar peduncle. The cerebellum in turn projects to the contralateral VL via the superior cerebellar peduncle, which decussates in the midbrain. The VL nucleus in turn projects to the motor cortex to complete the circuit. The cerebellum also receives unconscious proprioception from muscle spindles and Golgi tendon organs via the spinocerebellar and cuneocerebellar tracts. The cerebellum also projects to the ipsilateral vestibular nuclei, which give rise to the vestibulospinal tracts. The lateral vestibulospinal tract descends from the lateral vestibular nucleus to the spinal cord, where it facilitates ipsilateral extensor muscle tone of the trunk and extremities. As they descend, corticospinal fibers send collaterals to the ipsilateral red nucleus. The rubrospinal tract arises from the red nucleus and then immediately decussates and descends to facilitate flexor muscle tone, primarily in the upper extremities. The tectospinal tract arises from the superior colliculus, crosses in the dorsal tegmental decussation, and descends to influence muscles of the neck and upper back. It functions to move the head in response to external stimuli and to maintain head position in relation to the body position. The uncrossed pontine (medial) reticulospinal tract arises from the oral and caudal pontine reticular nuclei and facilitates extensor muscles, especially of the trunk and proximal extremities. The medullary (lateral) reticulospinal tract arises from the gigantocellular reticular nucleus and is primarily uncrossed but with a small crossed component. It travels just anterior to the rubrospinal tract. It inhibits antigravity muscles and is involved with autonomic functions.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


