Psychology and Biology of Memory
Andreas Meyer-Lindenberg
Terry E. Goldberg
Memory in psychiatric practice
Memory is the ability to store, retain, and retrieve information. This cognitive function plays a key role in psychiatry. Dementia and the amnesic disorders have memory dysfunction as a defining feature. Intrusive and recurrent emotional memories are one of the most distressing symptoms in post-traumatic stress disorder. Although not as obvious, problems with memory are also commonly revealed on testing in schizophrenia. Remembered episodes are often a focus in psychotherapy, as is the acquisition of new habits and response patterns. An ability to understand and assess memory is therefore important for the practising psychiatrist. In this chapter, basic neurobiological and psychological information
on memory will be reviewed. We have tried to cover a very broad field in a concise manner and give the interested reader a sense of the key memory systems and subsystems that are thought to be important for human information processing in health and in disease. We have emphasized the conceptual over the theoretical and key findings over the experimental details where possible. At times, we have not carefully separated the cognitive and neuroanatomical levels of analysis, both because they are sometimes almost inextricably bound and because it made our explanations clearer not to do so. Necessarily but not happily, we have omitted many important and active areas of investigation.
on memory will be reviewed. We have tried to cover a very broad field in a concise manner and give the interested reader a sense of the key memory systems and subsystems that are thought to be important for human information processing in health and in disease. We have emphasized the conceptual over the theoretical and key findings over the experimental details where possible. At times, we have not carefully separated the cognitive and neuroanatomical levels of analysis, both because they are sometimes almost inextricably bound and because it made our explanations clearer not to do so. Necessarily but not happily, we have omitted many important and active areas of investigation.
Forms of memory
One of the key discoveries of cognitive neuroscience is that ‘memory’ is not an unitary function, but consists of several forms that can be dissociated neurally and are differentially impacted by psychiatric disorders.(1) Several approaches can be taken to subdivide memory. One of the most straightforward is by the duration over which information is retained. In this way, ultrashort-term, short-term, and long-term memory can be distinguished. Ultrashort-term, also called sensoric or echoic/iconic memory, lasts from milliseconds to seconds and consists of a brief and modalityspecific retention of sensory information. For example, most people are able to ‘replay’ the auditory trace of the last second or so of a conversation, or briefly maintain a scene visualized after they close their eyes. In contradistinction, short-term memory has been shown to be relevant to a large number of psychiatric disorders. In short-term memory, information is briefly (over a period from seconds to minutes) held in mind, often through a process of rehearsal. A typical example is remembering a phone number from reading it to dialing without writing it down. A key feature of short-term memory is capacity limitation: most people are able to retain about seven items in short-term memory.(2) A specific form of short-term memory that has received considerable interest is working memory: the ability to hold information in mind that is necessary for a task at hand, but not present in the environment. This faculty is often regarded as a ‘mental workspace’ that is critical for information manipulation and goal-directed adaptive behavior, and the association of working memory with specific brain systems and psychiatric disorders has been widely studied. Rehearsal is also one important mechanism by which material is being transferred into long-term memory, which refers to the ability to retain information for time periods lasting from minutes up to the life span of the individual. This form of memory, which is also of major clinical importance, is not clearly capacity limited and is thought to depend on more enduring changes in neuronal structure and connectivity, raising the question of where in the brain these enduring memory traces, or ‘engrams’, are stored, how they are encoded for storage and how they are retrieved from it.
A second important subdivision is whether the content of memory can be consciously and intentionally retrieved (this is called explicit or declarative memory) or not (nondeclarative or implicit memory) (Fig. 2.5.3.1). Declarative memory is further subdivided in memory of facts and memory of events. Memory of events, which often includes recollection of temporal, spatial and emotional circumstances, is called episodic memory. Questions such as ‘What did you have for breakfast this morning?’ or ‘Where did you go to school?’ access episodic memory. Several features of episodic memory bear mention for their clinical relevance. For example, people can often say that they have seen a specific item before or that it ‘feels familiar’ without being able to recall the specifics of where and when (the episodic context). As discussed below, some evidence suggests that familiarity and recall may be supported by different brain regions. Intense feelings of familiarity without recall are experienced as déjà vu in a psychiatric context. People can also be convinced to remember events that did not, in fact, happen. These so-called false memories are also encountered in psychiatry. There has been much work on paradigms that produce false memories in normal individuals that address how they develop and how and why they might be successfully rejected. In one canonical account of episodic memory, the medial temporal lobe system (MTL) stores or indexes contextual markers that serve to bind feature information of a memory into an episodic configuration.
In contradistinction to episodic memory, memory of facts is not connected to specific experience. It is called semantic memory, and recollection of facts or vocabulary are examples of information that has become independent of episodic memory. Over time such material is thought to be stored in neocortex and can be retrieved without engagement of medial temporal lobe structures (MTL). Various models of the distinctions between semantic and episodic memory have proposed that while learning in the episodic system is rapid and can be based on a single trial or exposure, storage of information in the semantic system occurs slowly over time and only after multiple exposures or activations (3). Some accounts suggest that the MTL may also be involved in semantic memory, for example for separation or ‘decompression’ of stimuli previously learned as a unit.(4, 5) We have chosen not to review semantic memory in further detail because of space limitations and its complex overlap with psycholinguistics.
Whereas the subdivision of declarative memory is comparatively simple, implicit memory encompasses quite a heterogeneous group of functions that are supported by different brain systems. Among them are procedural memory, which refer to the gradual acquisition of sensorimotor, perceptual or cognitive skills through repeated exposure, priming (the facilitation of a response to an item if it was previously encountered), conditioning as well as various phenomena wherein a previously acquired response is gradually reduced or lost, such as extinction. Since the distinction between declarative and implicit depends on whether or not a memory process supports conscious recollection, clinicians must be careful not to confuse this with the properties of a given neuropsychological test; for example, a test of sentence completion requires conscious
production of words, but performance may strongly depend on implicit processes such as priming.
production of words, but performance may strongly depend on implicit processes such as priming.
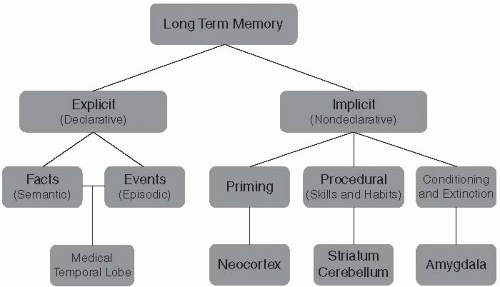 Fig. 2.5.3.1 Classification of memory and associated neural systems |
Although a taxonomy of memory systems is useful, many everyday tasks require functionality from several memory domains, and even relatively subtle changes in task demands may disrupt the balance among those cognitive systems. More refined analysis of learning tasks suggests that a variety of learning systems may mediate performance,(6) and that a given neural system participates in several forms of memory. For example, the hippocampal formation (HF), which is critical for episodic memory, is also thought to play a role in learning sequences so that indirect relations can be specified.(7) Thus, the HF becomes critical not during a>b, b>c, and c>d discriminations, but for the discrimination of the critical indirect b>d probe (e.g. if John is taller than Bill and Bill is taller than Mary and Mary is taller than Ellen, then Bill must be taller than Ellen).
Cellular and molecular mechanisms of memory
Memory is one of the most impressive examples of neural plasticity: the ability of the nervous system for enduring change triggered by external events. Arguably the best-studied cellular mechanisms underlying plasticity are long-term potentiation (LTP) and long-term plasticity (LTD), which mediate enduring changes on the level of the synapse.(8) Both have been best characterized in the hippocampus, one of the key structures for declarative memory.
By stimulating presynaptic fibres in the hippocampus (especially the CA1 section) with a brief pulse of high-frequency electric impulses, a long-lasting increase in responsiveness of the postsynaptic cells to low-frequency stimulation is reliably observed that can last for weeks (Fig. 2.5.3.2). This is called LTP. Initiation of this process depends on multiple second messenger mechanisms (Fig. 2.5.3.2). One of the best studied pathways starts with calcium influx into the presynapse through a glutamate receptor, NMDA, which activates further molecular cascades involving cAMP and protein kinases such as CamKII (other receptors, such as the glutamate receptors AMPA and mGluR, also play a role). LTP is then maintained by changes in gene transcription factors, such as CREB, and changed patterns of protein synthesis and phosphorylation, probably also dependent on protein kinase cascades. The time course of these processes can be used for a distinction between early phase LTP (the cellular signature of learning that occurs over seconds to minutes) and late phase LTP, which involves protein synthesis and occurs over minutes to hours and is thought to be critical for consolidation of new memoranda and would be linked to memory consolidation.(9,10)
LTD is a closely related process that is triggered when presynaptic stimulation is lower, causing less calcium influx (again through NMDA receptors) and preferential activation of calcineurin, a protein phosphatase. Together, LTP and LTD allow bidirectional enduring modulation of synaptic strength that could underlie formation and reversal of experience-dependent coupling in neural assemblies. Linking these synaptic changes to validated mechanisms underlying the complexities of human memory remains a challenge. However, the active field of neural network modeling has shown that, in principle, such changes in synaptic efficacy can produce efficient mechanisms to encode, store and retrieve information, a proposal first made by the neurophysiologist Donald Hebb.
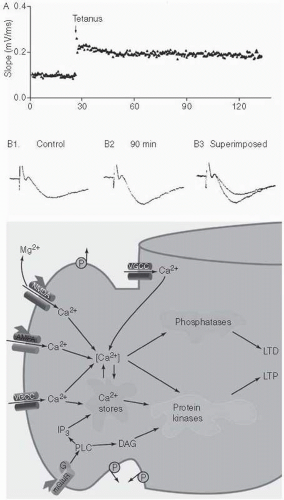 Fig. 2.5.3.2 Top: Long-term potentiation after a tetanic stimulation in the HF. Bottom: molecular mediators of long-term potentiation. |
Declarative/episodic memory
(a) Neural systems
Current evidence indicates that the HF and linked regions of the medial temporal lobe (MTL), in interactions with parts of the prefrontal cortex, play a critical role in the encoding and retrieval of episodic memories, whereas engrams are stored in neocortex(11) (Fig. 2.5.3.3). Interactions of the HF with amygdala are important for emotional memories. The HF consists of the hippocampus proper, the entorhinal cortex, which provides the main port of entry for connections with the cortex, and the adjacent perirhinal and parahippocampal cortices, which interact with the entorhinal cortex and in turn receive projections from all other neocortical areas, with parietal, dorsal occipital and prefrontal regions primarily projecting to parahippocampal gyrus and temporal cortex to
perirhinal cortex. In this way, the HF is bidirectionally connected with the rest of the brain. Although pertinent observations were already made at the end of the 19th century by Bechterew, the importance of the hippocampal formation for episodic memory was dramatically shown in 1953 by the case of patient H.M., in whom the HF and amygdala were resected bilaterally as a treatment for drug-resistant epilepsy(12) (Fig. 2.5.3.4). This led to complete and enduring anterograde amnesia (inability to form new episodic memories). In addition, he has some degree of retrograde amnesia (i.e. an inability to retrieve episodic information stored before the operation), while his working and procedural memory, as well as priming, is unimpaired. A similar pattern of memory impairment is observed in Wernicke encephalopathy/Korsakov’s syndrome or other neurological processes impacting on diencephalic structures, including the medial thalamus, mamillary body and the fornix, which project to the HF. This indicates that these structures may be viewed as a system that is critical for assigning a spatiotemporal (episodic) context. In rodents, a major role of the HF is indeed to function as a neural map of the environment;(13) it is controversial to what degree this applies to humans. Neuroimaging studies confirm that activation of the HF is observed during successful encoding and retrieval of episodic information.(14)
perirhinal cortex. In this way, the HF is bidirectionally connected with the rest of the brain. Although pertinent observations were already made at the end of the 19th century by Bechterew, the importance of the hippocampal formation for episodic memory was dramatically shown in 1953 by the case of patient H.M., in whom the HF and amygdala were resected bilaterally as a treatment for drug-resistant epilepsy(12) (Fig. 2.5.3.4). This led to complete and enduring anterograde amnesia (inability to form new episodic memories). In addition, he has some degree of retrograde amnesia (i.e. an inability to retrieve episodic information stored before the operation), while his working and procedural memory, as well as priming, is unimpaired. A similar pattern of memory impairment is observed in Wernicke encephalopathy/Korsakov’s syndrome or other neurological processes impacting on diencephalic structures, including the medial thalamus, mamillary body and the fornix, which project to the HF. This indicates that these structures may be viewed as a system that is critical for assigning a spatiotemporal (episodic) context. In rodents, a major role of the HF is indeed to function as a neural map of the environment;(13) it is controversial to what degree this applies to humans. Neuroimaging studies confirm that activation of the HF is observed during successful encoding and retrieval of episodic information.(14)
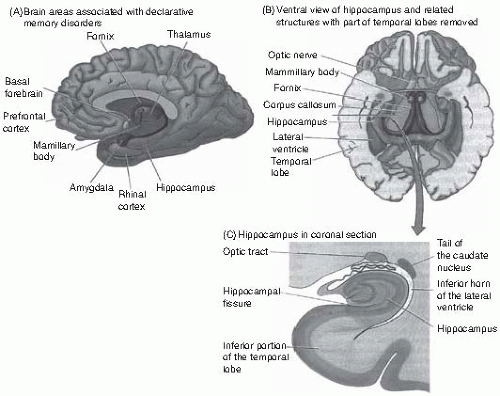 Fig. 2.5.3.3 (A) brain regions associated with dysfunction in episodic memory. (B) the hippocampal formation, view from below. (C) coronal section through the hippocampal formation. |
A neurocognitive model system for examining the computational role of MTL subsystems involved in episodic memory may help the reader gain a sense of the interplay among various subprocessing modules.(15) In this model, the MTL binds memories and their instance-specific context and then stores their code for later retrieval. The model described here is based on studies showing that the bulk of hippocampal (HC) cortical input is segregated over two pathways. One of these may convey spatial information; the other may convey information regarding items and objects. The two streams are interconnected at various levels within entorhinal cortex, which likely contributes to the integration of cortical inputs into a representation of their co-occurrence. The hippocampus proper may quickly associate a code to the conjunction of cortical inputs, such that similar entorhinal patterns come to be separated via their associated hippocampal patterns. These hippocampal patterns are thus not directly associated with individual features, but serve to separate the large number of overlapping entorhinal patterns, which is important to ensure that retrieval will be unambiguous. The representational overlap in entorhinal cortex, combined with the pattern separation system in the hippocampus proper, enormously increases the storage capacity of the memory store and allows accurate recall of episodic memories. In this model, retrieval in this memory system can be sampled using cues, consisting of partial input patterns; for instance, part of a context representation from a previously experienced episode. Initially, such a cue may activate only part of an associated entorhinal pattern, but if the set of activated entorhinal nodes sufficiently resembles a stored representation, their combined firing will tend to activate associated hippocampal nodes, through previously strengthened connections with these nodes. The hippocampal nodes, in turn, will recruit missing nodes of the entorhinal representation. This pattern completion process will reinstate the original pattern in the entorhinal layer, which, in turn, can reinstate associated information in the input layers, namely, item representations that have been experienced in that particular context (feature extraction). Thus, all features of an episode can be recalled, even when only part of the input layers is cued.
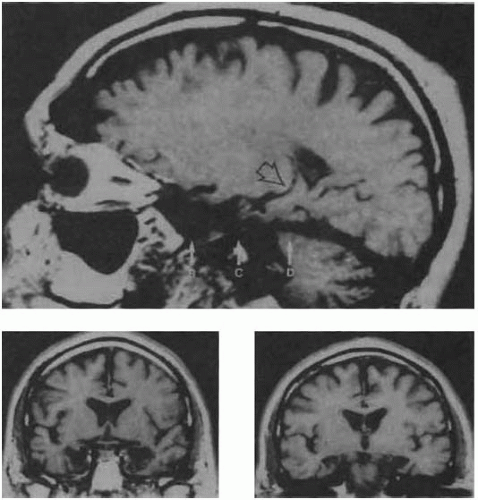 Fig. 2.5.3.4 Hippocampal damage in patient H.M., shown on a sagittal (top) and two coronal slices (below) of the patient’s MRI. |
The model was shown to be sensitive to various types of simulated lesions including reductions of nodes (‘neurones’), addition of noise to the system, and perhaps most relevant for the modeling of schizophrenia, marked reductions in connectivity between the various modules. At the network level, this reduced connectivity led to compromised cross-association of episodic features (i.e. item and context) and a superimposed, mild reduction of pattern separation in the system. The latter malfunction made some
patterns irretrievable, affecting all memory tasks including recognition, albeit to a mild degree. The cross-association problem also attenuated ‘searching’ of the memory store, particularly with single-source cues. This preferentially affected tasks with a large retrieval demand, such as free recall. From an information processing standpoint, what appeared to be disproportionate failures in retrieval were due to compromised encoding.
patterns irretrievable, affecting all memory tasks including recognition, albeit to a mild degree. The cross-association problem also attenuated ‘searching’ of the memory store, particularly with single-source cues. This preferentially affected tasks with a large retrieval demand, such as free recall. From an information processing standpoint, what appeared to be disproportionate failures in retrieval were due to compromised encoding.
Recent work has indicated that recollection is predominantely mediated by the hippocampus proper, whereas familiarity has been linked to perirhinal cortex.(16) While this distinction is supported by functional neuroimaging studies of healthy individuals (e.g. Eldridge et al.[17]), studies of patients with circumscribed lesions of the hippocampus have nevertheless found severe familiarity-based recognition memory impairments.(18) These results do not necessarily contradict a dual process distinction, but could rather suggest that MTL regions comprise an integrated network that supports both processes.
There has been much interest in the precise role of episodic memory systems in the formation of associations between items that are entering memory. Several new studies suggest that the hippocampus is engaged preferentially when inter-item associations are formed in memory. While there have been several compelling accounts that specific subsystems in the MTL complex play different roles in single item encoding and associative encoding, the distinction between the two may be relative, not absolute.(11, 19)
Neuroimaging and lesion studies have suggested that interactions between the HF and the amygdala are relevant for emotional memories, especially if these have a fearful or aversive character.(20) Neuroimaging has also demonstrated that encoding and especially retrieval is associated with activation of the lateral prefrontal cortex, as well as with increased functional interactions of these regions with the HF that are supported by anatomical tracts such as the uncinate fascicle. In some studies, left prefrontal cortex is differentially more involved than right in encoding information into episodic memory, whereas right prefrontal cortex is differentially more involved than left in episodic memory retrieval.(21) Compared to encoding and retrieval, the evidence is much less clear with regard to the storage of the engrams themselves. Cases such as H.M. and the clinical picture of Korsakov’s syndrome show that the HF and diencephalic structures cannot be the store, since most episodic and semantic memories laid down before the onset of illness are spared. Current evidence suggests that the neocortex is the ultimate store of memories and that engrams reside in regions that are also specialized in processing stimuli to which they pertain. For example, circumscribed cortical lesions can result in categoryspecific impairments in retrieving object information, the so-called anomias, which have been described for classes such as people, tools, or living things, and neuroimaging studies show that similar regions are differentially activated during naming of these object classes. In each case, engrams are assumed to be stored in a distributed pattern of synaptic connections over a large group of neurones. It is an open question how the interaction of the HF and cortex accomplishes the encoding, and retrieval of information from these neural assemblies (but see above for a model). On the molecular level, glutamatergic neurotransmission is crucial to support the LTP mechanisms that support the neural plasticity essential for memory formation. In addition, acetylcholine is a neurotransmitter associated with declarative memory function since it is known that muscarinic receptor blockade impairs episodic memory and degeneration of cholinergic neurones in the basal nucleus (of Meynert) is a prominent finding in Alzheimer’s disease. It is likely that cholinergic mechanisms act on declarative memory by modulating glutamate-dependent LTP and LTD in MTL regions.
Related posts:


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


