Psychology of Attention
Elizabeth Coulthard
Masud Husain
Introduction
Attention is generally taken to be the process by which people are able to concentrate on certain information or processes, while ignoring other events. It appears to be a fundamental attribute of human brain processing, although difficult to pin down in terms of mechanism. Psychologists have attempted to fractionate attention in many different ways, using ingenious behavioural paradigms. In this section we, too, will consider different aspects of attention: selective, phasic and sustained, divided and executive control of attention. However, it would be fair to say that all these aspects of attention do not normally operate in isolation. Instead they interact, and deficiencies in one aspect of attention, for example, in a patient population, often to do not occur in isolation. Functional imaging and lesion studies of attention have proliferated in recent years, attempting to place a neurobiological framework to these varied processes. In general, these studies also tend to confirm the view that attention is likely an emergent property of widespread brain networks, with a special emphasis on frontal and parietal regions of the human brain (Fig. 2.5.2.1). In this discussion we illustrate several aspects of attention with examples particularly from literature on visual attention, which is the most widely studied area, but it should be appreciated that many of the concepts discussed here extend to other domains. In fact, there is a good deal of evidence to suggest that several aspects of attention operate at a supra- or cross-modal level allowing integration of information from different sources.
Recent studies suggest there are two fronto-parietal networks: (Fig. 2.5.2.1) a dorsal parieto-frontal network involving the superior parietal lobe (SPL) and dorsal frontal regions such as the frontal eye field (FEF); and a ventral network involving the inferior parietal lobe (IPL), temporoparietal junction (TPJ) and inferior frontal gyrus (IFG). In addition, dorsomedial frontal areas, including the anterior cingulate cortex (ACC) and pre-supplementary area (pre-SMA) may play a key role in flexible control of attention for strategic behaviour.
Selective attention
Selective attention refers to the processes involved in selecting relevant information and filtering out irrelevant items from the vast array of information we are exposed to. The brain has limited capacity: it simply cannot process everything it is exposed to. Nor would it be sensible for it do so because the majority of sensory input to which it is exposed is not behaviourally relevant. Therefore there is a need for mechanisms to select the
most behaviourally significant and important material, and dispose of trivial, unimportant information. Such selection may occur ‘bottom-up’, driven by competition between sensory inputs, or ‘top-down’, guided by the goals that an individual might have at any moment in time.(1)
most behaviourally significant and important material, and dispose of trivial, unimportant information. Such selection may occur ‘bottom-up’, driven by competition between sensory inputs, or ‘top-down’, guided by the goals that an individual might have at any moment in time.(1)
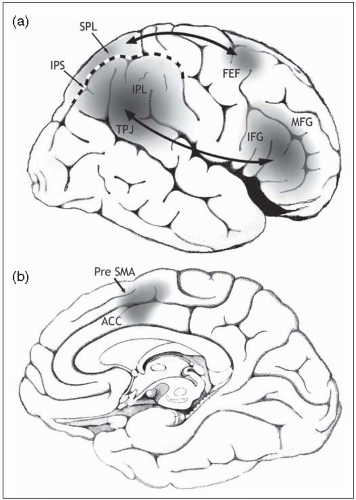 Fig. 2.5.2.1 Lateral (a) and medial (b) regions in the right hemisphere involved in attention. (IPS intraparietal sulcus; MFG, middle frontal gyrus.) |
Thus if a ball is unexpectedly hurled towards an observer, his attention is likely to be captured—‘bottom-up’—by the sensory input of a projectile moving at high velocity towards his head. In this case, selection has been driven by the most perceptually salient item in the external environment. ‘Top-down’ attention mechanisms, on the other hand, concern selection that is biased by internal goals. For example, consider the processes involved in looking for a friend in a busy, crowded train station. Here the selection process is driven by the features you are searching for: your friend’s hair colour, facial features, height all play a role in guiding this search. Importantly, under these circumstances, even perceptually very salient items may be filtered out—or not attended—if they are irrelevant to the task.
But when does selection occur? Is it early or late in the processing of sensory information? This is an issue that dominated attention research for many years. Some investigators proposed that selection occurs early, directly after analysis of the physical characteristics or features of sensory stimuli, but before they are fully identified. According to this view, unattended information receives little or no further processing from this point on. By contrast, others argued that all stimuli are analysed up to the point that they are identified. Selection occurs only after this, late in the processing stream. So items that are eventually ignored or unattended, i.e. those that are not selected, actually receive considerable processing before they are discarded. Note that this late selection model allows for the possibility that items which are eventually ignored may nevertheless be processed to a deep level. Thus, even though they may not be attended to, they have the potential to influence our actions subliminally. Most researchers would now agree that there is good evidence for both early and late selection systems in the human brain. In fact, whether selection occurs early or late is likely to be influenced by the specific demands of the task.
Two highly influential experimental paradigms used to study selective attention in healthy humans and patients have both focused on spatial attention in the visual system. Posner first developed a spatial cueing task in which subjects view a display consisting of a central cross on which they are asked to maintain fixation throughout the trial. On either side of the cross, there are two square boxes. Participants are instructed to ignore a cue which consists of transient illumination of either the left or right box. At varying intervals after the cue, a target stimulus (an asterisk) appears in either left or right box and subjects are required to press a response button as quickly as possible. In 80 per cent of trials the cue is ‘valid’ in that it occurred at the location of the subsequent target. However, in the remaining 20 per cent of trials the cue is ‘invalid’, appearing in the box opposite that in which the target subsequently appeared. In healthy volunteers, reaction times to targets appearing where valid cues appear are significantly shorter than when invalid cues are presented.
This critical finding suggests that attention can be spatially localized like a beam or ‘spotlight’. Moreover, attention can be captured by the abrupt onset of the cue so that visual processing is selectively oriented towards it, thereby improving responses if a target subsequently appears there. On invalid trials, Posner argued, attention would first have to disengage from the invalidly cued location and then shift to the correct location before engaging it. Note that such shifts occur covertly in the absence of overt eye movements; they represent shifts of visual processing from one location to another across a representation of space in the brain. Subsequent studies have shown that the orienting of attention to a spatial location not only appears to speed up detection of a visual stimulus, as measured by reaction times, but also can improve discrimination of items from non-targets. The neurophysiological mechanisms underlying such boosting of performance are currently the subject of intense scrutiny.(2)
The second experimental task that has proven to be extremely important in selective attention research is the visual search paradigm developed by Treisman. In this task, subjects have to find a target shape embedded among distractors. In simple so-called ‘feature search’, a target may be defined by a unique feature, e.g. a red circle among green circles is defined uniquely by its colour and therefore ‘pops-out’ among the distractors. Treisman has considered that such feature searches can occur pre-attentively in parallel across the visual scene, without the need for a spotlight of visual attention. She argued that attention needs to be deployed in more complex tasks where a target may share one or more features with distractors, e.g. finding a red circle among green circles and
red squares. In this case, a single feature (colour or shape) is not sufficient to define the target. Instead, the visual system has to find the unique conjunction of red colour and circular shape, and the target does not pop-out to the observer. Such ‘conjunction searches’, Treisman argued, requires the spotlight of attention to shift serially from one location to the next, inspecting each item in turn. In her model, spatial attention acts to bind or glue together features occupying the same location in space, e.g. the colour, form, luminance, and other attributes that belong to an object at one location in space.
red squares. In this case, a single feature (colour or shape) is not sufficient to define the target. Instead, the visual system has to find the unique conjunction of red colour and circular shape, and the target does not pop-out to the observer. Such ‘conjunction searches’, Treisman argued, requires the spotlight of attention to shift serially from one location to the next, inspecting each item in turn. In her model, spatial attention acts to bind or glue together features occupying the same location in space, e.g. the colour, form, luminance, and other attributes that belong to an object at one location in space.
Both the Posner cueing task and Triesman’s conjunction task have been used in neuroimaging studies of healthy volunteers and patients with focal lesions on the brain.(3) The results of such studies suggest that regions within the parietal and frontal cortex play a critical role in deploying spatial attention (Fig. 2.5.2.1). In general, dorsal regions of this parieto-frontal network have been implicated in the spatial shifts of processing attention that occupy such a key role in the models of selective attention developed by Posner and Treisman.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


