Somatosensory Systems: Introduction
Input from the somatosensory systems informs the organism about events impinging on it. Sensation can be divided into four types: superficial, deep, visceral, and special. Superficial sensation is concerned with touch, pain, temperature, and two-point discrimination. Deep sensation includes muscle and joint position sense (proprioception), deep muscle pain, and vibration sense. Visceral sensations are relayed by autonomic afferent fibers and include hunger, nausea, and visceral pain (see Chapter 20). The special senses—smell, vision, hearing, taste, and equilibrium—are conveyed by cranial nerves (see Chapters 8, 15, 16, and 17). In addition, nociceptive sensation or pain-signaling serves to warn the organism when there is contact with noxious or potentially damaging elements in the environment, or when tissue is damaged.
Receptors
Receptors are specialized cells for detecting particular changes in the environment. Exteroceptors include receptors affected mainly by the external environment: Meissner’s corpuscles, Merkel’s corpuscles, and hair cells for touch; Krause’s end-bulbs for cold; Ruffini’s corpuscles for warmth; and free nerve endings for pain (Fig 14–1). Receptors are not absolutely specific for a given sensation; strong stimuli can cause various sensations, even pain, even though the inciting stimuli are not necessarily painful. Proprioceptors receive impulses mainly from pacinian corpuscles, joint receptors, muscle spindles, and Golgi tendon organs. Painful stimuli are detected at the free endings of nerve fibers.
Each efferent fiber from a receptor relays stimuli that originate in a receptive field and gives rise to a component of an afferent sensory system. Each individual receptor fires either completely or not at all when stimulated. The greater the intensity of a stimulus, the more end-organs that are stimulated, the higher the rate of discharge is, and the longer the duration of effect is. Adaptation denotes the diminution in rate of discharge of some receptors on repeated or continuous stimulation of constant intensity; the sensation of sitting in a chair or walking on even ground is suppressed.
Connections
A chain of three long neurons and a number of interneurons conducts stimuli from the receptor or free ending to the somatosensory cortex (Figs 14–1, 14–2, and 14–3).
The cell body of a first-order neuron lies in a dorsal root ganglion or a somatic afferent ganglion (eg, trigeminal ganglion) of cranial nerves.
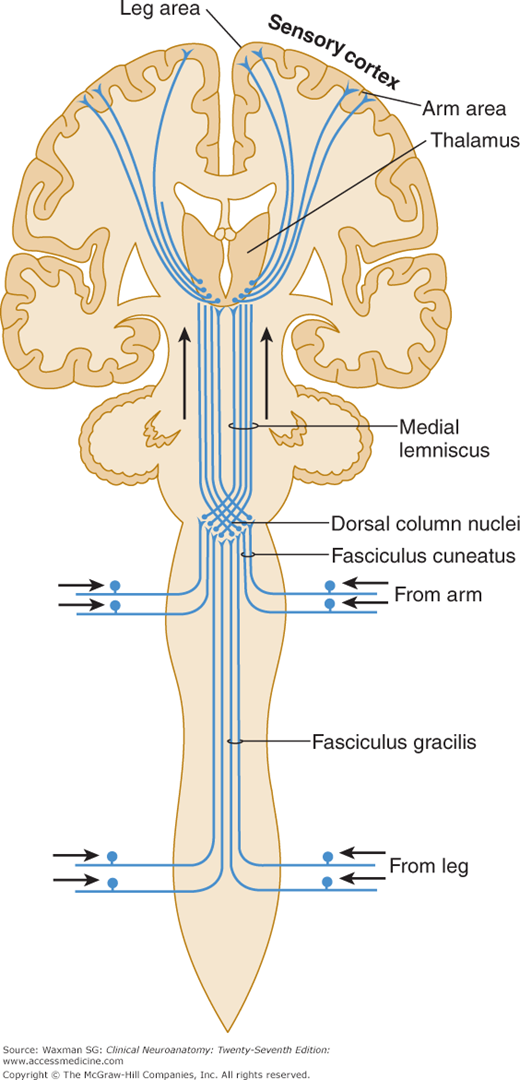
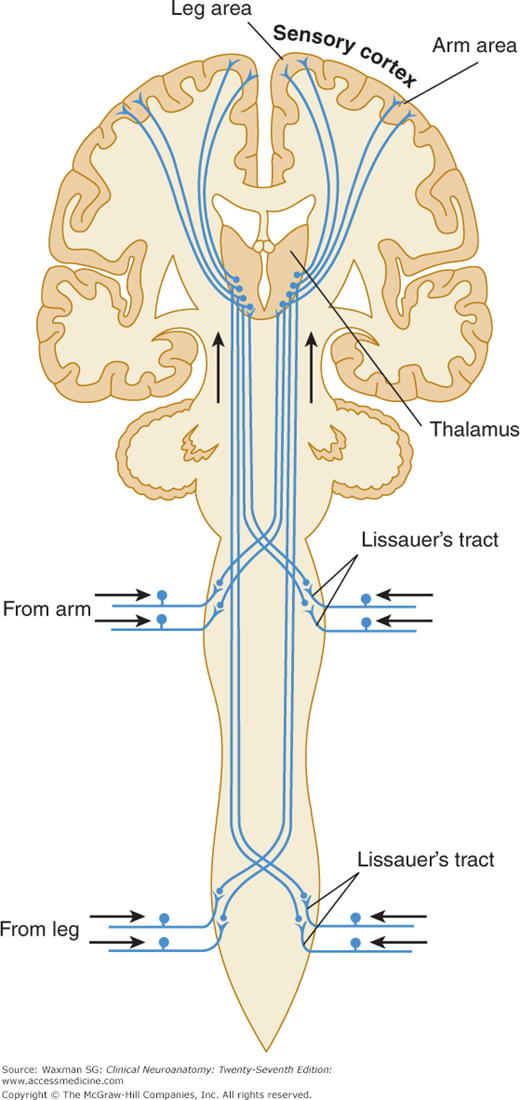
The cell body of a second-order neuron lies within the neuraxis (spinal cord or brain stem; examples are provided by the dorsal column nuclei, ie, the gracile and cuneate nuclei, and by neurons within the dorsal horn of the spinal cord). Axons of these cells usually decussate and terminate in the thalamus.
Clinical Correlations
Interruption in the course of first- and second-order neurons produces characteristic sensory deficits, which can be especially apparent when they involve sensitive areas such as the face or fingertips. An example is provided by sensory loss in the territory innervated by a particular nerve or spinal root when this nerve or root is injured.
Sensory Pathways
Multiple neurons from the same type of receptor often form a bundle (tract), creating a sensory pathway. Sensory pathways ascending in the spinal cord are described in Chapter 5; their continuation within the brain stem is discussed in Chapter 7. The main sensory areas in the cortex are described in Chapter 10.
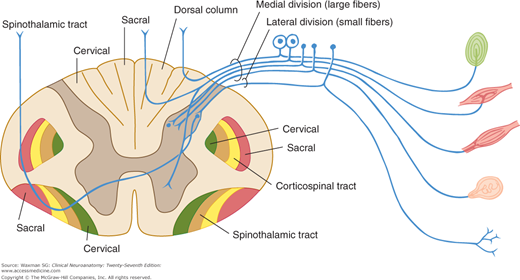
One major system—the lemniscal (dorsal column) system (see Fig 14–2)—carries touch, joint sensation, two-point discrimination, and vibratory sense from receptors to the cortex. The other important system—the ventrolateral system—relays impulses concerning nociceptive stimuli (pain, crude touch) or changes in skin temperature (see Fig 14–3). Significant anatomic and functional differences characterize these two pathways: the size of the receptive field, nerve fiber diameter, course in the spinal cord, and function (Table 14–1). Each system is characterized by somatotopic distribution, with convergence in the thalamus (ventroposterior complex) and cerebral cortex (the sensory projection areas; see Figs 10–13 and 10–15), where there is a map-like representation of the body surface. The sensory trigeminal fibers contribute to both the lemniscal and the ventrolateral systems and provide the input from the face and mucosal membranes (see Figs 7–8 and 8–12).
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


