Fig. 5.1
The stress-diathesis model of suicidal behavior
Although cumulative evidence in several domains supports a stress-diathesis model of suicidal behavior, there are also conflicting results that highlight its complexities. Psychosocial stressors such as poor social adjustment, job loss, or declining health have been consistently associated with an increased risk of suicidal behavior (Blackmore et al. 2008). However, the results of prospective studies examining the effect of life events on suicidal behavior have not been conclusive (Oquendo et al. 2013). When examining longitudinally the determinants of suicide attempts, Oquendo et al. (2013) found that health- and work-related life events, as well as major depressive episodes, increased by 13-fold the risk of a subsequent suicide attempt among depressed subjects, provided that they did not have comorbid borderline personality disorder. They also found that predisposing features such as female gender or suicidal cognitions were related to the risk and timing of the suicidal act. Yet, among depressed subjects with borderline personality disorder, the effect of life events or major depressive episodes on suicidal behavior appeared to be buffered. Borderline personality disorder, a condition particularly predisposed to suicidal behavior because of its association with pessimism, impulsivity, and early trauma, seemed paradoxically to cope better with negative life events. The authors hypothesized that a lower threshold for stress could explain these findings; “normal” events could be as important as life events for borderline patients.
On the other hand, clinical studies have shown important differences between suicide attempters and non-attempters independently of related mental disorders. Suicide attempters present higher levels of impulsive aggression, hopelessness, and more frequent familial history of suicidal behavior, personal history of childhood adversities, and head injury. All of these factors are part of a diathesis for future suicidal behaviors (Krakowski and Czobor 2004; Lopez-Castroman et al. 2012; Oquendo et al. 2004; Perroud et al. 2008). However, although most subjects with mental disorders never attempt suicide, psychopathology is probably the main risk factor for suicidal behavior, present in 90 % of completed suicides and 80 % of suicide attempts (Nock et al. 2010). In global rates there is a particular contribution of affective disorders to suicidal behavior because of their high prevalence, but anxiety disorders, psychotic disorders, eating disorders, and substance use disorders (particularly use of alcohol or nicotine), as well as borderline personality disorder, also contribute significantly (Bolton and Robinson 2010).
There is also convincing evidence on the influence of biological factors both as predisposing and precipitating agents of suicidal behavior. The most consistent biological finding in suicidal behavior is the implication of the serotonin system, the dysfunction of which represents a constitutional risk factor. The largely replicated findings of an association between low cerebrospinal fluid (CSF) levels of serotonin and suicidal behavior, independently of psychiatric disorder, gave a substantial weight to the existence of a specific biological suicidal vulnerability. A large number of studies using different indicators of serotonergic dysfunction suggest that this dysfunction may act as a vulnerability trait of suicidal behavior (Mann 2003). On the other hand, abnormal stress responses such as hyperactivity of the hypothalamic-pituitary-adrenal axis (HPA), evaluated with the dexamethasone suppression test, are linked with suicidal behavior (Coryell and Schlesser 2001). Lastly, a decrease in cholesterol and brain-derived neurotrophic factor (BDNF) levels, measured in blood serum or plasma, have also been associated with impaired brain plasticity among individuals with suicidal behavior (Lee and Kim 2011).
Mental or “psychic” pain is a concept that describes pervasive negative affect that is experienced by the sufferer as pain (Orbach et al. 2003). Such affect can fluctuate somewhat widely in intensity, and some authors have pointed to mental pain as an effective potential trigger of suicidal acts. Mental pain also seems to increase the propensity to suicidal ideation (Olié et al. 2010), and can be assessed and targeted for prevention (Mee et al. 2011). Interestingly, mental pain might increase the risk of severe suicidal behaviors especially when accompanied by difficulties with social communication and loneliness (Gvion et al. 2014).
The study of the notes left by suicide completers and psychological autopsy studies indicate that suicide victims were nearly always suffering from a psychiatric disorder and submitted to environmental stressors. However, environmental stressors and psychiatric disorders are necessary but non-sufficient conditions for attempting suicide. Long-term prospective studies reveal that suicide attempters are at high risk of committing suicide in their lifetime (about 13 % will) (Tidemalm et al. 2008). The personal history of suicide attempt is the main risk factor for subsequent suicidal behavior, indicating the existence of a vulnerability to suicide. Additionally, individuals suffering from a mental disorder are at particularly high risk during the first years of illness. This suggests that suicidal vulnerability is more easily expressed when subjects are exposed for the first time to major stresses represented by a psychiatric condition. As an example, in a study of patients with unipolar depression, rates of suicidal behavior were 74.4 % for the first episode, 18.8 % for the second one and 6.5 % for subsequent ones (Mann et al. 1999).
Prospective studies bring more evidence supporting the stress-diathesis model. A cohort of adult affective patients after a major depression episode that was followed prospectively for 2 years in New York showed an additive effect of the three most powerful predictors of future suicidal acts (history of suicide attempt, subjective severity of depression, and cigarette smoking) and also of another two predisposing factors: pessimism and impulsive-aggression (Oquendo et al. 2004). Of note, another prospective study by the same group demonstrated that clinical predictors of suicidal acts vary between genders (Oquendo et al. 2007). In a 21-year longitudinal study of a birth cohort of 1,265 New Zealand young adults, those who developed major depression had increased rates of suicidal ideation and suicide attempt. Again, suicidal ideation or suicide attempts appeared in a minority of depressed young adults. Thus, additional factors seemed to modify the vulnerability (or resilience) to suicidal responses, including: familial history of suicide, childhood abuse, personality features, peer affiliations, and school performance (Fergusson et al. 2003).
5.3 Current Directions: Genetics, Neuroimaging and Neuropsychology
Genetic factors play a significant role in suicidal behaviors, and can interact with stressful events to increase the risk of suicidal behavior. In an innovative study, Caspi (2003) revealed that a variant of the serotonin transporter gene was associated with increased risk of suicidal ideation after significant life events. This finding was later replicated in abused depressed patients (Shinozaki et al. 2013), psychiatric inpatients with a history of suicide attempts (Gibb et al. 2006), and substance-dependent patients (Roy et al. 2007). Apart from monoaminergic systems, other gene x environment (GxE) interactions involving glutamatergic, GABAergic and corticotrophin related genes are being investigated, as well as genes involved in neurotrophic processes (particularly the gene encoding BDNF) or in inflammatory processes (Mandelli and Serretti 2013; Perroud et al. 2008). Independently of psychopathology, concordance rates of suicidal behavior ranging from 6 to 35 % have been reported in studies examining monozygotic twins (Pedersen and Fiske 2010). Of note, although the transmission of suicidal behavior across generations seems to be independent of Axis-I disorders, it could be mediated by intermediate phenotypes (endophenotypes), like impulsive aggression or hopelessness (Courtet et al. 2011).
Few studies have connected stress and suicide through neuroimaging. Jollant et al. (2008) compared the neural correlates of implicit emotional processing in euthymic males with a history of depression according to history of suicide attempt. Men with a past history of suicide attempt showed increased sensitivity to anger reflecting others’ disapproval, were more likely to act on negative emotions, and paid less attention to mildly positive stimuli. These results support the hypothesis of a predisposition for suicidal acts in negative circumstances (Jollant et al. 2008). More recently, Reisch et al. (2010) used fMRI through the presentation of autobiographical scripts to reactivate the memories of a recent suicide attempt in females. Three conditions were examined: mental pain, suicide action, and neutral activity. The recollection of mental pain was associated with decreased prefrontal activity compared to the recollection of suicide action, which showed an increased activity in the frontal cortex. To explain their findings the authors hypothesized that goal-directed suicidal behavior would be undertaken to reduce the mental pain, which would act as a traumatic stressor.
The pursuit of neurocognitive markers of suicide vulnerability constitutes also a very productive area of research. Suicidal behavior may be viewed as a consequence of an altered decision process when facing stressful situations. Proof of such alteration has been found in several studies mostly using the Iowa Gambling Task among adolescent (Bridge et al. 2012) or adult populations (Jollant et al. 2005, 2010) of suicide attempters in which attempters appear to ignore past experiences when performing a reward/punishment task (Dombrovski et al. 2010). Importantly, the use of computerized association tests is a promising area of research that could be used to identify individuals at risk in stressful situations (Cha et al. 2010). Moreover, ecological momentary assessments through portable devices can be used to obtain real-life longitudinal data of affective variability and suicidal ideation in response to environmental stressors (Palmier-Claus et al. 2012).
5.4 Life Is More Complex: Vulnerability Contributes to Stress
On the one hand, alterations in decision-making, a putative suicidal endophenotype involved in the ability of the individual to make choices in daily life, are correlated to the occurrence of problematic affective relationships (Jollant et al. 2007), threatening individual’s social bounds. On the other hand, according to the theory of attachment (Bowlby 1977), the perception of being rejected or neglected, and by extension being abused in childhood or in adulthood, leads to rejection sensitivity and the perception of being unwanted in general (Ehnvall et al. 2008). Being excluded or rejected leads to reactive behaviors, which could be dysfunctional: aggressive, antisocial, or self-injurious behaviors (Williams 2007). To be impulsive/aggressive or to be less prone to approach others due to pessimism/hopelessness exacerbates the risk of social isolation. Social isolation and interpersonal difficulties, both psychosocial stresses relevant to precipitate suicidal acts, may be partially due to suicidal vulnerability factors. Interestingly, contacting people by telephone one month after being discharged from an emergency department for deliberate self-poisoning may help reduce the number of suicide re-attempts over 1 year, independently of the identification of subjects at risk or the implementation of a crisis intervention (Vaiva et al. 2006). This highlights the very powerful effect of perceived social isolation.
Suicidal vulnerability may lead the subjects to consider non-stressful environmental events as stressors because of hypersensitivity to psychological pain or social exclusion. Indeed, using functional MRI, ventromedial prefrontal and cingulate regions associated with suicidal vulnerability (Jollant et al. 2010, 2008) have also been involved in social rejection/distress (Eisenberger et al. 2003) and psychological pain perception (van Heeringen et al. 2010). Thus, the suicidal act relies on a stress-vulnerability model, complicated by the role of vulnerability to emerging stress.
5.5 Connecting Vulnerability-Stress with Neurodevelopment
The interaction of disturbances during development with genetically modulated temperaments may explain the triggering effects of later life stressors through personality and coping styles. In other words, vulnerability traits determined by alterations in neurodevelopment and biological substrates interact dynamically with acute stressors enabling suicidal behavior.
5.5.1 Prenatal and Perinatal Risk Factors
Neurodevelopment starts during pregnancy. Different research teams have described a “birth season” effect in suicide, which may be independent of any associated psychopathology. The largest study to date examined 6.5 millions persons and 80,000 suicides in Hungary and reported that suicide risk was associated with a spring or summer birth: being born in July increased the risk by 14 % compared with a birth in December (Döme et al. 2010).
Several hypotheses have been proposed to explain these surprising findings. Brain insults by infectious agents during development could convey vulnerability for suicidal behavior when interacting with particular genetic conditions. Seasonal variations of serotonin metabolism, inducing decreases in CSF serotonin levels, could also be associated with impulsivity and, secondarily, with an augmentation of suicidal risk (Chotai et al. 2006). Finally, melatonin rates and circadian rhythms vary depending on the season of birth among healthy new-born and the vesperal chronotype is associated with impulsivity, which could lead to suicidal behavior (Chotai 2005). Interestingly, subjects born in risk periods such as spring present more often the “S” allele of the serotonin transporter that is associated with suicidal vulnerability (Gonda et al. 2012).
Two very large cohorts followed prospectively for 30 years in Sweden and Scotland support the influence of perinatal factors on suicidal risk. Future suicide attempts, studied only in the Swedish cohort, were increased in subjects with small birth length, multiparity, low sociocultural level, and young age of the mother (Mittendorfer-Rutz et al. 2004). The risk of completed suicide was associated with low birth weight and young maternal age in both studies, but an association with multiparity and unemployment of the parents was additionally found in the Scottish cohort (Riordan et al. 2006). On the other hand, postnatal growth could also play a significant role. A weak weight gain in the first year after birth was associated with a higher risk of suicide for 15,000 English subjects born between 1911 and 1930. This finding was independent of birth weight, socio-economic level, and the type of diet of the new-born (Barker et al. 1995). Finally, very preterm birth, childhood maltreatment, and personality traits seem to have additive effects that influence the age at onset of the first suicide attempt (Blasco-Fontecilla et al. 2013).
5.5.2 The Role of Childhood Maltreatment
Childhood maltreatment is strongly associated with suicidal behavior in adulthood (Short and Nemeroff, this volume). It is also a prevalent life event that can be reported by a majority of suicide attempters in some populations (Lopez-Castroman et al. 2012). Childhood abuse interacts with family and genetic factors in increasing the risk of early onset suicidal behavior, and higher impulsive aggressive traits (Lopez-Castroman et al. 2012, 2014; Slama et al. 2009). The mechanisms that link childhood maltreatment and suicidal behavior remain unclear. Literature on neurodevelopment has demonstrated that early trauma affects cognitive and affective processes causing less intellectual performance, memory impairments, alterations in executive functions, and emotional dysregulation (Pechtel and Pizzagalli 2010). These impairments may confer an increased risk of adult psychopathology and facilitate the onset of suicidal behaviors. Several authors have also proposed impulsive aggression as a mediating factor between childhood abuse and suicidal behavior. Although causal relations cannot be explored without prospective studies, high impulsive aggression in abused patients is associated with more severe suicidal behaviors (Lopez-Castroman et al. 2014).
5.5.3 Biological Embedding
Biologic transfer mechanisms, particularly changes in gene expression, might explain the impact of negative life experiences, such as childhood abuse, in the multifactorial process of suicidal behavior (See Dwevedi, this volume). The mechanisms that allow early experiences to modulate brain development and determine vulnerability or resilience seem to implicate epigenetic factors. Epigenetic modifications lead to the repression or activation of genetic expression, and have been identified in several genes (related with glucocorticoid receptors, BDNF, arginine vasopressin system, and corticotropin-releasing hormone) in connection with early environmental changes in animal models (review in: Turecki et al. 2012).
Although most of these studies examined post-natal epigenetic regulation, stress during pregnancy may also induce epigenetic changes. Studies with rodents highlight the importance of the timing of epigenetic alterations. Stress during early pregnancy seem to allow an epigenetic reprogramming of the rat genome causing long-term modifications on the HPA axis. The type of stress is also important, minimal stressors could be associated with resilience, while massive or prolonged stress would lead to developmental frailty.
The exact role of vulnerability genes in suicidal behavior warrants further research. For instance, how the “s” allele of the serotonin transporter increases risk? This is paradoxal because this allelic variant is associated with a diminished recapture of serotonin, the mode of action of serotoninergic antidepressants that have shown effectiveness in the prevention of suicide reattempts. Could this genetic polymorphism act through neurodevelopmental means? Studies with knockout mice support this hypothesis. If the mutation is viable, it would induce large modifications with homeostatic consequences on neurodevelopment (Lesch and Mössner 2006). Additionally, the early role of serotonin and its receptors in the course of prenatal development, as well as its influence on the morphogenesis of the brain, suggests that serotonin influences the development and brain maturation of mammals before taking its role as a neurotransmitter. Besides, there seems to be a common genetic background between neurodevelopmental alterations and suicidal behavior (Perroud et al. 2008).
5.5.4 Toward a Neurodevelopmental Model of Suicidal Behavior
The neurodevelopment hypothesis was initially used to explain the pathophysiology of schizophrenia. But being nonspecific, it is applicable to other mental disorders, including suicidal behavior. Prospective cohort studies show that impulsive aggression in childhood remains stable until adulthood and is associated with an elevated risk of suicidal behaviors (Pihlakoski et al. 2006). Thus, Caspi et al. (1996) described an increased risk of antisocial personality or depression in adulthood for children of 3 years of age showing lack of self control or behavioral inhibition, respectively. In both cases, the risk of suicidal behaviors was increased (Fig. 5.2).
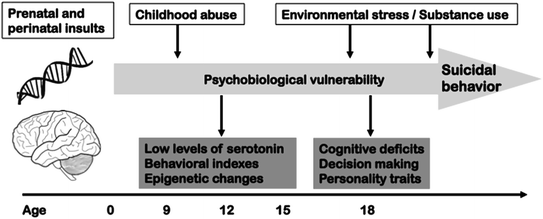
Fig. 5.2
A developmental model of suicidal behavior
At a biological level, serotoninergic abnormalities leading to suicidal vulnerability have also been observed among adolescent subjects (Tyano et al. 2006). During adolescence, environmental stressors may exacerbate imbalances in the function of limbic and ventral prefrontal areas, leading to suicidal behavior, a risk that may be mediated by genetic factors (Casey et al. 2010). Biological data for young children is scarce, but points in the same direction (Clarke et al. 1999). The stability of impulsive-aggression traits connected to serotoninergic systems (Oxenstierna et al. 1986) favors the idea of an early onset of psychobiological disturbances that would be later expressed as auto- or hetero- aggressive behaviors. Studies of epidemiological and molecular genetics report similar findings in childhood and adolescence (Dwivedi and Zalsman 2012).
There are also some neuropsychological indexes that support a neurodevelopmental model of suicidal behavior. A large Swedish prospective study found a clear association between low intelligence and later suicide risk, independently of socioeconomic level or mental disorders (Gunnell et al. 2005). Intellectual deficits could be the consequence of alterations in early brain development and explain the risk of suicidal behavior in adulthood. As mentioned before, decision-making constitutes also a cognitive trait of suicide vulnerability that is modulated by the interaction of sexual abuse in childhood and a genotypic variant of the CRH-R1 gene (Guillaume et al. 2013). Recent data suggest that anomalies in decision-making might appear already in adolescence (Oldershaw et al. 2009).
Related posts:
 Traumatic Brain Injury (TBI) and Post Traumatic Stress Disorder (PTSD) as Risk Factors for Suicidal Thoughts and Behaviors
Genetic Risk Factors for Suicidal Behavior
Experimental Pharmacologic Approaches for the Reduction of Suicidal Ideation and Behavior
Psychotherapeutic Treatment Approaches for Suicidal Individuals
Traumatic Brain Injury (TBI) and Post Traumatic Stress Disorder (PTSD) as Risk Factors for Suicidal Thoughts and Behaviors
Genetic Risk Factors for Suicidal Behavior
Experimental Pharmacologic Approaches for the Reduction of Suicidal Ideation and Behavior
Psychotherapeutic Treatment Approaches for Suicidal Individuals
 Animal Models of Risk Factors for Suicidal Ideation and Behaviour
Animal Models of Risk Factors for Suicidal Ideation and Behaviour
 Treatment Emergent Suicidal Ideation and Behavior
Treatment Emergent Suicidal Ideation and Behavior
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


