Fig. 2.1
Constantin von Economo (1876–1931), Professor of Psychiatry and Neurology at the University of Vienna, left, and Georg N. Koskinas (1885–1975), former Assistant at the Psychiatric and Neurological Clinic of the University of Athens, right. For his discovery of encephalitis lethargica, von Economo was nominated three times for the Nobel Prize in Physiology or Medicine between 1926 and 1932. Koskinas returned to Greece in 1927; after an unsuccessful application for the chair of Neurology at the University of Athens, he devoted himself to private practice in the suburb of Kifisia (Triarhou 2005) (Photo credits: Bildarchiv und Grafiksammlung der Österreichischen Nationalbibliothek, Vienna (Economo); Helios Encyclopedic Lexicon, Athens (Koskinas). Used by permission and protected by copyright law. Copying, redistribution or retransmission without the author’s express written permission is prohibited)
With his landmark monograph, Brodmann (1909) defined 44 cortical cytoarchitectonic areas in the human brain (and a total of 52 areas in the primate brain overall). He studied cortical cytoarchitecture in numerous mammals, from the hedgehog, with its unusually large archipallium, to primates and humans, and introduced the terms homogenetic and heterogenetic formations to denote two different basic cortical patterns with either the typical six layers or lacking the six-layer stage, respectively (Garey 2006; Zilles and Amunts 2010).
Vogt and Vogt (1919) laid the foundations of myeloarchitectonics (the architecture of fiber pathways) and defined the structural features of allocortex, proisocortex and isocortex; they also analyzed the differences between paleocortical, archicortical, and neocortical regions (Vogt and Vogt 1919; Vogt 1927; Zilles 2004; Zilles and Amunts 2012).
Von Economo commenced his work on cortical cytoarchitectonics in 1912, and Koskinas joined him in 1919. Their Atlas and Text Volume were published in 1925, and included 150 new discoveries (Koskinas 1931). Von Economo and Koskinas (1925, 2008) defined 107 area modifications, and more than 60 area transitions (von Economo 2009), virtually raising the “resolution” of our cortical cytoarchitectonic register, compared to Brodmann’s data, by a factor of four.
In subsequent decades, by combining cytoarchitectonics with myeloarchitectonics, Sanides (1962, 1964) placed emphasis on transitions or gradations that accompany “streams” of neocortical regions coming from paleocortical and archicortical sources (Pandya and Sanides 1973), while Vogt and Vogt (1919) had already spoken of “areal gradations”.
Comprehensive tables correlating the 107 cortical areas defined by von Economo and Koskinas with the Brodmann areas can be found in a previous review article (Triarhou 2007b) and in the English edition of the Atlas (von Economo and Koskinas 2008).
The work of von Economo and Koskinas represents a gigantic intellectual and technical effort (van Bogaert and Théodoridès 1979). Their attempt to bring the existing knowledge into a more orderly pattern was emphatically acknowledged by von Bonin (1950) and Bailey and von Bonin (1951).
Spyridon Dontas (1878–1958), Professor of Physiology and Pharmacology at the University of Athens and President of the Academy of Athens, had remarked in 1926 upon meeting Koskinas: “The work of von Economo and Koskinas is monumental and constitutes a milestone of science, charting new paths for understanding the brain from an anatomical, physiological and pathological viewpoint. It stands as the first comprehensive reference on the architecture of the adult human cerebrum and will persevere as a perpetual scientific testimony” (Triarhou 2012).
The brain map and the systematic area naming by von Economo and Koskinas have regrettably not passed into widespread general use. However, it is clear that they brought together concepts and ideas of cortical organization and structure that had been developing over the preceding 30 years and which remain with us in the present era of cortical research; moreover, they introduced original terms and, by applying in a systematic manner nomenclatures derived from other authors and themselves, they codified the language that we use to describe the cortex to this day, essentially providing the first “ontology” of the cerebral cortex (Jones 2010).
2.2 Method
At the outset of their studies, von Economo and Koskinas devised an entire system of new methods to overcome the existing obstacles and difficulties, from the autopsy to the photographic documentation (Koskinas 1926, 1931; von Economo 2009; von Economo and Koskinas 2008). The following are some of the introduced innovations.
2.2.1 Sectioning
Instead of the widely adopted method of sectioning the whole brain serially, perpendicular to its fronto-occipital axis, von Economo and Koskinas obtained tissue sections always perpendicular to the axis of each gyrus or sulcus and in directions corresponding to their convoluted pattern (Figs. 2.2 and 2.3). They arrived at that idea by considering that, in order to be able to compare the various brain areas cytoarchitectonically, sections had to have a consistent orientation relative to the gyral surface, insofar as only then could the breadth of the entire cerebral cortex and of each cortical layer as well be represented in the sections in a precise way.
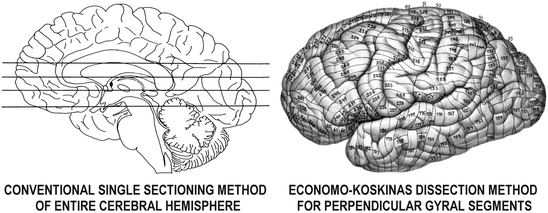
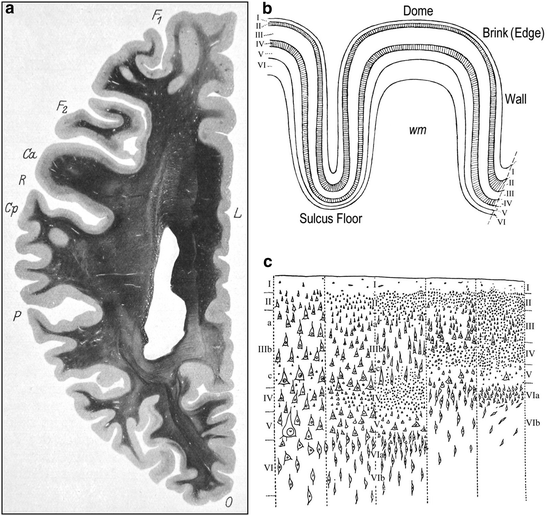
Fig. 2.2
The difference between the widely used method of obtaining whole single sections of the cerebral hemispheres, left, and the method devised by von Economo and Koskinas (1925, 2008) for dissecting each hemisphere into 250–350 tissue blocks, 4 mm in thickness, always perpendicular to the axis of each gyral or sulcal segment, right; hatched areas indicate “cancelled” tissue
Fig. 2.3
(a) In obtaining sections of the entire cerebral hemisphere through conventional sectioning techniques, the real variations in layer thickness and cellular architecture cannot be studied consistently. The horizontal section through the left human cerebral hemisphere depicts such sizeable regional differences in cortical thickness and the random orientation of the gyri (von Economo and Koskinas 1925). Weigert method. F 1 and F 2, superior and middle frontal gyrus; Ca, precentral gyrus; R, central sulcus; Cp, postcentral gyrus, P, parietal lobe; O, occipital lobe; L, limbic gyrus. (b) A schematic drawing that depicts the varying thickness of the six cortical layers (I through VI) at the level of the dome, brink (edge), wall and valley (sulcus floor) in a cortical gyrus. The two granular layers (external and internal) are hatched; wm, subcortical white matter (von Economo 2009). (c) The five fundamental structural types of isocortex: 1, agranular; 2, frontal; 3, parietal; 4, polar; 5, granulous or koniocortex (von Economo 1925, 1929, 2009; von Economo and Koskinas 1925, 2008)
2.2.2 Staining
The staining of the preparations was perfected such that a uniform tone was achieved not only of the single sections, but of all the series of sections into which each brain had been divided. That was mandated by the need, firstly, to define gradual differences of histological elements in neighboring areas of the cerebral cortex, and secondly, to achieve consistent photographic registrations.
2.2.3 Specimen Depiction
Most of the previous histological studies on cortical cytoarhitecture depicted their results schematically, and therefore subjectively. Instead of schematic drawings, and aiming at an exact documentation of the specimens, with all the relationships of the diverse neurons, von Economo and Koskinas used photography, which is the most objective testimony regarding form, size and arrangement, and turned to branches of science such as advanced optics and photochemistry.
The stained cortical sections were photographed using Carl Zeiss Planar lenses, which are special macro objectives with a considerably larger field than the common microscopy objectives, especially valuable for large area objects under relatively large magnifications. Planar lenses are used without an eyepiece. Additional details on technique can be found in my historical notes on Koskinas (Triarhou 2005) and von Economo (Triarhou 2006).
The depth of field that von Economo and Koskinas achieved in their photomicrographs, as well as the clarity and detail with which individual neurons can be visualized is remarkable. Their plates probably still represent the most comprehensive set of high resolution images of cortical histology ever assembled (Jones 2008).
2.3 General Part
The “General Part” of the Text Volume (von Economo and Koskinas 1925) covers introductory concepts on the cerebral cortex and its nerve cells, the structure and the development of the cortical layers, the composition and the meaning of the cortical laminar structure, the definition of cortical areas, and methodological issues (Fig. 2.4).

Fig. 2.4
Schematic map of the lateral (convex) facies of the hemispheric surface and three microphotographic plates from the Atlas of von Economo and Koskinas (2008), shown as examples: Plate XV – Magnocellular intermedio-agranular frontal area FCB m (Broca’s area) at the foot of the inferior frontal gyrus, anterior wall. Plate XXX – Triangular granular frontal area FD γ in the inferior frontal gyrus, wall of the notch of pars triangularis (incisura capi). Plate XCIV – Supratemporal area granulosa TC in first gyrus of Heschl (primary auditory cortex), middle, dome, with the typical “rain shower formation” (Regenschauerformation). The detailed descriptions of the normal histological structure of the cerebral cortex depicted in the 112 microphotographic plates of the Atlas were explained in the accompanying Text Volume (von Economo and Koskinas 1925). The printing size of the original plates was 40 × 40 cm at a magnification of × 100, therefore covering a 4.0 × 4.0 mm true cortical area
Brodmann (1909) grouped his 44 human cortical areas as 4 postcentral, 2 precentral, 8 frontal, 4 parietal, 3 occipital, 10 temporal, 6 cingulate, 3 retrosplenial, and 4 hippocampal.
Von Economo and Koskinas (1925, 2008) divided the cortex into seven lobes, which they denoted by their initials. The lobes were further subdivided into regions: the frontal lobe (F) into prerolandic, anterior (prefrontal) and orbitomedial (orbitomedial) regions; the superior limbic lobe (L) into anterior, posterior and retrosplenial regions; the parietal lobe (P) into postcentral (anterior parietal), superior, inferior and basal regions; and the temporal lobe (T) into supratemporal, temporal proper, fusiform and temporopolar regions. The insular (I) and occipital (O) lobes were not subdivided. The inferior limbic lobe consists of the hippocampus (H). For cytoarchitectonic area designations, they did not continue Brodmann’s system of random numbers, but instead used letter codes, consisting of a Roman capital letter (the initial of the lobe), followed by a calligraphic capital to note the sequence of a gyrus within a lobe (e.g. FB means the second gyrus of the frontal lobe), and a Latin or Greek subscript for characteristic microscopic features (e.g. m = magnocellular, p = parvicellular, γ = gigantopyramidal).
Von Economo and Koskinas (1925, 2008) defined five fundamental “supercategories” of structural cortical types (agranular, frontal, parietal, polar and granulous) (Fig. 2.3c), further arranged into 54 ground, 76 variant and 107 cytoarchitectonic modification areas, plus more than 60 transition areas (von Economo 1925, 2009; von Economo and Horn 1930). Topographically, the 107 modification areas of von Economo and Koskinas are grouped into 35 frontal, 13 superior limbic, 6 insular, 18 parietal, 7 occipital, 14 temporal, and 14 inferior limbic or hippocampal (Figs. 2.5 and 2.6). Of the 107 modifications, 22 are allocortical, 22 heterotypic isocortical, and 63 homotypic isocortical. Von Economo and Koskinas (1925, 2008) separately analyzed the dome, edge, wall and floor of each cortical gyrus (Fig. 2.3b).
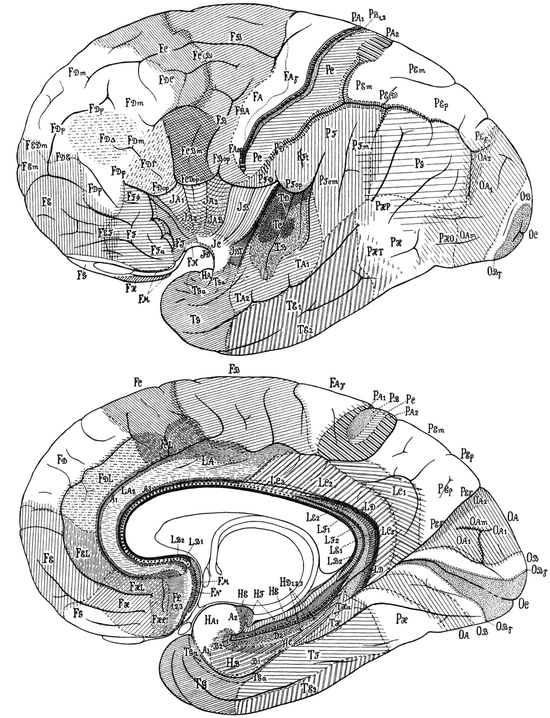
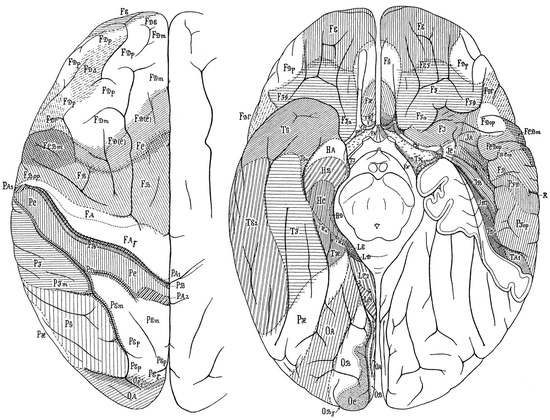
Fig. 2.5
Cytoarchitectonic maps of von Economo and Koskinas, showing cortical modification areas in the convex and median hemispheric facies of the human brain
Fig. 2.6
Cytoarchitectonic maps of von Economo and Koskinas for the dorsal and ventral hemispheric surface
For certain cortical areas with a granular appearance of their cells in most layers, especially of gyral walls, associated primarily with sensory functions, von Economo and Koskinas (1923, 1925) introduced the term koniocortex to denote their dusty appearance.
Von Economo and Koskinas (1925, 2008) regularly saw a special type in a small band in sublayer IIIc at the boundary between any koniocortex (or sensory isocortex) and the ordinary surrounding isocortex in sensory parietal, occipital and temporal areas. Such zones contain giant pyramidal cells. They called these margin regions with magnocellular characteristics parasensory zones.
Another crucial discovery was that of the large, spindle-shaped bipolar projection neurons in the inferior ganglionic layer (Vb) of the dome of the transverse insular gyrus, which are now called “von Economo neurons” (Watson et al. 2006) – although a more succinct term might be “von Economo-Koskinas neurons”. The detailed morphology of these rod cells (Stäbchenzellen) and corkscrew cells (Korkzieherzellen) was documented by von Economo and Koskinas (1925) in cingulate (anterior) limbic and frontoinsular areas.
2.4 Special Part
The following text is a selection of some ideas discoursed in previous reviews (Triarhou 2007a, b) and in the new English editions of the Atlas (von Economo and Koskinas 2008) and of von Economo’s shorter textbook of cortical cytoarchitectonics (von Economo 2009).
2.4.1 Frontal Lobe
Broca’s motor speech area FCB m in the inferior frontal gyrus was considered as a particular human characteristic by von Economo and Koskinas (1925), as well as by Brodmann (1909). The surface area of the pars opercularis of the inferior frontal gyrus is characterized by a distinct type of cortex, distinguishable from the posteriorly lying premotor cortex in area FB in the precentral gyrus; it continues rostrally as area FD γ (Fig. 2.4).
Anteriorly one finds portions of areas FD and FE, which are rich in granule cells. Lesions in the prefrontal region result in disturbances of attention, psychomotor activity, will and emotivity. Von Economo (2009) termed such higher mental functions, localized in the frontal regions of the brain, “the active part of the psychic personality”. Area FA γ resembles the frontal core area more closely, with the consequence that a large part of area FA belongs to nonprimary motor cortex.
Area FF partly corresponds to the orbitofrontal proisocortex of the monkey that lies intercalated between the caudal orbitofrontal isocortex rostrally, and the orbitofrontal peripaleocortex caudally. Area FF a in the human brain probably corresponds to the granular isocortex in the anterior part of the orbital surface of the frontal lobe in the macaque. Areas FH and FHL correspond to the paralimbic dysgranular isocortex on the ventromedial surface of the prefrontal cortex in the macaque, which lies intercalated between the frontopolar granular isocortex rostrally and the orbitomesial archicortical proisocortex of the straight gyrus caudally. Area FJ appears to correspond to peripaleocortex in the inferior part of the transverse gyrus of the insula, and to the orbitofrontal peripaleocortex in the monkey (de Olmos 1990). The area FF lies rostrally and ventrally to area FD γ .
Von Economo and Koskinas (1925) mark transitional types of cortex in their maps, beyond the 107 “standard” modifications; such transitions comprise the areas FBA, FC(B), FCD op, FDC, FDE, FED m , FEF and FE m . Areas FBA, FDC, FDE and FEF denote transition forms (e.g. FBA marks the transition of area FB into FA, FDC the transition of FD into FC, and so on). The designation FC(B) implies a part of area FC with an admixture of the type of the neighboring area FB, whereas the subscript m in the areas FED m and FE m signifies cellular variations with magnocellular features. Area FCD op is a transitional opercular variant between areas FC op and FD op.
2.4.2 Parietal Lobe
Brodmann (1909) defined four areas (1, 2, 3, 43) in the postcentral region, whereas von Economo and Koskinas six (PA 1, PA 2, PB 1, PB 2, PC and PD). In the parietal region, Brodmann defined four areas (5, 7, 39, 40), and von Economo and Koskinas nine (PED, PE m , PE p , PE γ , PF, PF t , PF op, PF cm and PG). The basal parietal region PH most likely belongs to the visual cortex and includes the functionally defined areas V4 and V5 (Zilles and Palomero-Gallagher 2001). The proposed subdivisions of the anterior parietal cortex by von Economo and Koskinas are still in use.
Area PA is located in the depths of the central sulcus. Area PB is “sensory koniocortex”, located on the caudal bank of the central sulcus. Concerning the primary somatosensory cortical areas and the subdivision of the posterior parietal lobe into a superior and an inferior lobule, the accepted terminology of von Economo and Koskinas forms the basis for modern cytoarchitectonic analyses and experiments in primates (Zilles 2004). Brodmann did not delineate any transition zones in the posterior parietal lobe, whereas von Economo and Koskinas marked such transition zones between the areas PE, PF, PG, PH and OA, in agreement with the observations of Eidelberg and Galaburda (1984).
Related posts:
 Where Matters: New Approaches to Brain Analysis
Where Matters: New Approaches to Brain Analysis
 Visualizing Myeloarchitecture In Vivo with Magnetic Resonance Imaging in Common Marmosets (Callithrix jacchus)
Visualizing Myeloarchitecture In Vivo with Magnetic Resonance Imaging in Common Marmosets (Callithrix jacchus)
 High-Field Magnetic Resonance Mapping of the Border Between Primary Motor (Area 4) and Somatosensory (Area 3a) Cortex in Ex-Vivo and In-Vivo Human Brains
High-Field Magnetic Resonance Mapping of the Border Between Primary Motor (Area 4) and Somatosensory (Area 3a) Cortex in Ex-Vivo and In-Vivo Human Brains
 MRI Methods for In-Vivo Cortical Parcellation
MRI Methods for In-Vivo Cortical Parcellation
 The Myeloarchitectonic Studies on the Human Cerebral Cortex of the Vogt-Vogt School, and Their Significance for the Interpretation of Functional Neuroimaging Data
The Myeloarchitectonic Studies on the Human Cerebral Cortex of the Vogt-Vogt School, and Their Significance for the Interpretation of Functional Neuroimaging Data
 Estimating the Location of Brodmann Areas from Cortical Folding Patterns Using Histology and Ex Vivo MRI
Estimating the Location of Brodmann Areas from Cortical Folding Patterns Using Histology and Ex Vivo MRI
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


