Figure 15-1. Development of the diencephalon. Lateral (A) and midsagittal (B) views of the forebrain at about 8 to 9 weeks of gestational age. The cross-sectional views (C, D) are taken from the planes shown in B and emphasize diencephalic structures.
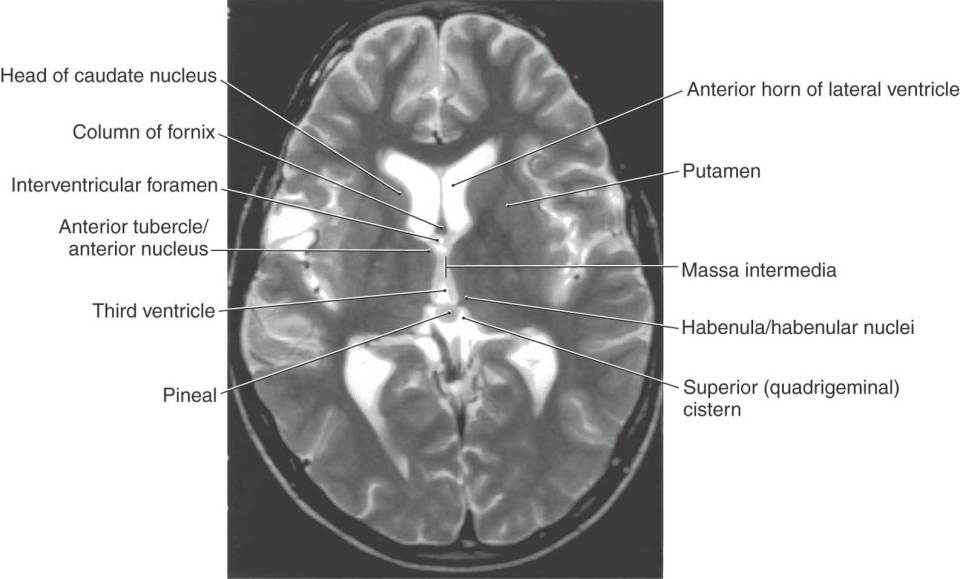
Figure 15-2. Axial T2-weighted magnetic resonance image showing the interventricular foramen, the massa intermedia, the area of the habenula, and the pineal in the superior cistern. The interventricular foramen is the space (containing a small portion of choroid plexus) located between the column of the fornix and the anterior tubercle of the thalamus. The major nuclei of the dorsal thalamus in an axial plane are shown in Figure 15-10.

Figure 15-3. Midsagittal view showing the locations of circumventricular organs.

Figure 15-4. Midsagittal view of the diencephalon and closely related structures. This is a drawing of the specimen shown in Figure 15-6.
Just anterior to the habenular region, the roof plate epithelium and adjacent pia mater give rise to the choroid plexus of the third ventricle, which, in the adult, remains suspended from the roof of this space (Fig. 15-1B-D). This choroid plexus is continuous through the interventricular foramina with that of the lateral ventricles. Elsewhere, in locations around the perimeter of the third ventricle, specialized patches of ependyma lie on the midline and form unpaired structures called the circumventricular organs. These structures include the subfornical organ, the organum vasculosum of the lamina terminalis, the subcommissural organ, and the pineal gland (Fig. 15-3). These cellular regions are characterized by the presence of fenestrated capillaries, which implies an absence of the blood-brain barrier. These structures are thought to release metabolites and neuropeptides into the cerebrospinal fluid or into the cerebrovascular system.
The development of the pituitary gland during the third week is linked to that of the diencephalon (Fig. 15-1B, C). A downward extension of the floor of the third ventricle, the infundibulum, meets the Rathke pouch, an upward outpocketing of the stomodeum, the primitive oral cavity. By the end of the second month, the Rathke pouch loses its connection with the developing oral cavity but maintains its attachment to the infundibulum. As development continues, the Rathke pouch gives rise to the anterior lobe (adenohypophysis) and pars intermedia of the pituitary gland, whereas the infundibulum differentiates into the posterior lobe of the pituitary gland, or neurohypophysis (Fig. 15-1B; see Chapter 30). A craniopharyngioma (Rathke pouch tumor) can arise from a portion of the Rathke pouch that fails to undergo proper migration and apposition to the infundibulum. These tumors mimic lesions of the pituitary gland and may cause visual problems, diabetes insipidus, and increased intracranial pressure.
BASIC ORGANIZATION
The junction between the diencephalon and midbrain lies along a line extending from the posterior commissure to the caudal edge of the mammillary body on the medial aspect of the hemisphere (Fig. 15-4). On the surface of the hemisphere, this interface is represented by a line starting at the caudal aspect of the mammillary body, extending anterolaterally over the edge of the crus cerebri, and following the caudal edge of the optic tract (Fig. 15-5). The boundary between the diencephalon and surrounding telencephalon is less distinct and is represented laterally by the internal capsule and rostrally by the interventricular foramen, lamina terminalis, and optic chiasm (Fig. 15-4).
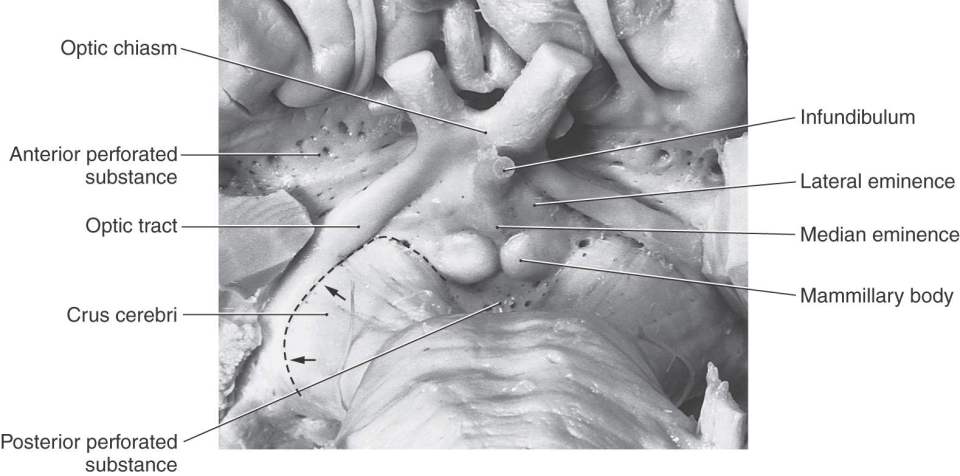
Figure 15-5. Anterior (ventral) view of the hemisphere emphasizing diencephalic structures visible on the surface and showing the diencephalic-mesencephalic interface as represented by the caudal edge of the optic tract (arrows).
The cavity of the diencephalon, the third ventricle, is a narrow, vertically oriented midline space located between the paired dorsal thalami and hypothalami of the two sides (Figs. 15-6 and 15-7). In addition to its connections with the lateral ventricles and the cerebral aqueduct, the third ventricle has small evaginations or recesses associated with the optic chiasm (supraoptic recess), the infundibulum (infundibular recess), and the pineal gland (pineal and suprapineal recesses) (Figs. 15-1 and 15-4).
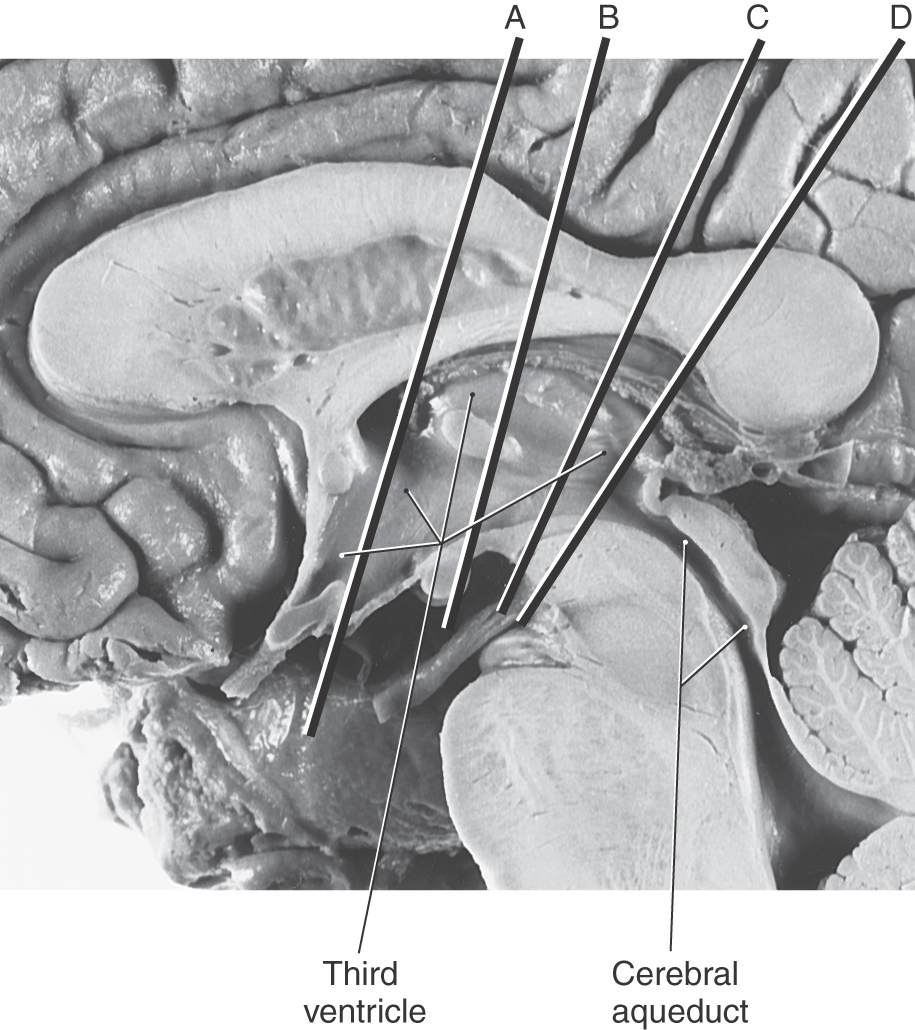
Figure 15-6. Midsagittal view of the diencephalon. This view correlates with the drawing in Figure 15-3. Lines A through D indicate the planes of the stained sections in Figure 15-7.
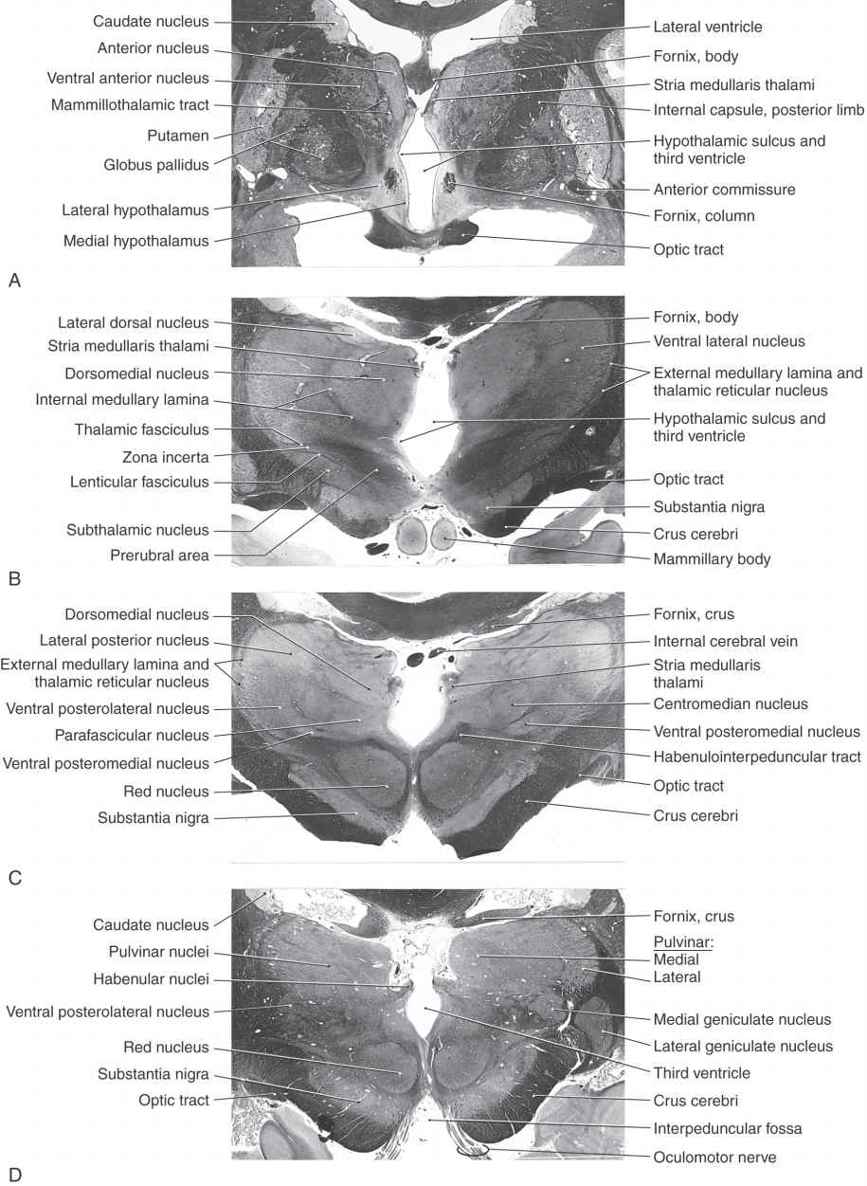
Figure 15-7. Four levels of the forebrain from rostral (A) to caudal (D) showing the internal structure of the hemisphere with emphasis on the diencephalon. These levels correlate with those shown in Figure 15-6 and with the planes represented in the exploded view in Figure 15-10. Weil stain.
All four diencephalic subdivisions can be approximated in a midsagittal section of the forebrain (Figs. 15-4 and 15-6). The dorsal thalamus is located superior to the hypothalamic sulcus and extends from the interventricular foramen caudally to the level of the splenium of the corpus callosum. The hypothalamus lies inferior to the hypothalamic sulcus and is bordered rostrally by the lamina terminalis and caudally by a line that extends from the posterior aspect of the mammillary body superiorly to intersect with the hypothalamic sulcus. The only diencephalic structures visible on the inferior surface of the hemisphere are those related to the hypothalamus, including the optic chiasm, infundibulum, medial and lateral eminences, and mammillary bodies (Fig. 15-5). The ventral thalamus (subthalamus) does not border on the ventricle; rather, it occupies a position caudal to the hypothalamus, rostral to the diencephalon-midbrain junction, and lateral to the midline (Figs. 15-4 and 15-7B). Epithalamic structures are located posteriorly and caudally, in close apposition to the posterior commissure, and include the pineal gland, the habenular nuclei, and the main afferent bundle of these nuclei, the stria medullaris thalami.
DORSAL THALAMUS (THALAMUS)
The dorsal thalamus (or thalamus) (Figs. 15-4 and 15-7 to 15-9) is a massive collection of neuronal cell groups that participate in a widely diverse array of functions involving motor, sensory, and limbic systems. It receives a variety of ascending inputs and projects, via thalamocortical fibers to various cortical areas or gyri, and receives reciprocal connections, via corticothalamic fibers, from those cortical targets to which it sends projections. As a result, the thalamus is often regarded as the functional “gateway” to the cerebral cortex.

Figure 15-8. Magnetic resonance images of the cerebral hemisphere in the coronal plane from rostral (A) to caudal (D). Although many structures are clearly seen, emphasis in the labeling is placed on diencephalic structures. All images are T1 weighted.
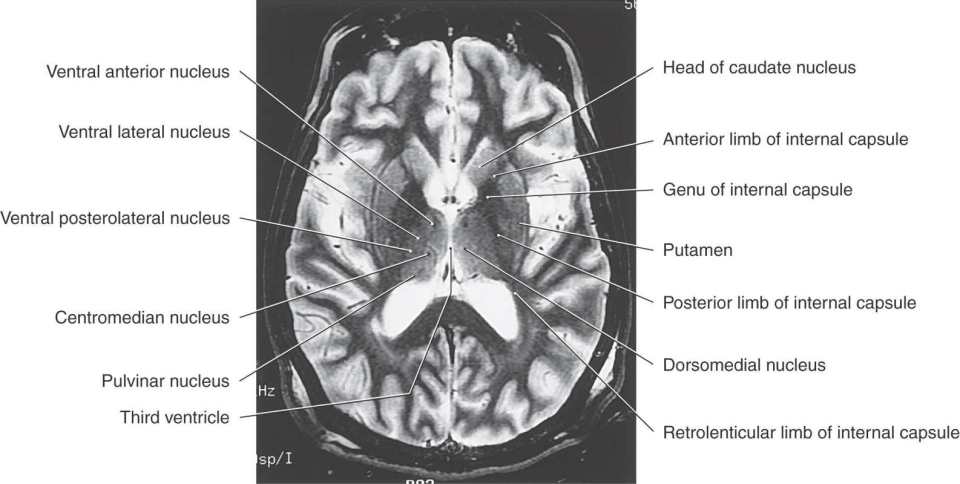
Figure 15-9. T2-weighted magnetic resonance image of the cerebral hemisphere in the axial plane. Emphasis in labeling is on diencephalic structures. Compare with Figure 15-12.
The thalamus is covered on its lateral aspect by a layer of myelinated axons, the external medullary lamina, which includes fibers that enter or leave the subcortical white matter (Fig. 15-7B, C). Within the external medullary lamina are clusters of neurons that form the thalamic reticular nucleus. The medial surface of the thalamus borders the third ventricle, and the external medullary lamina and thalamic reticular nucleus blend with the thalamic fasciculus and zona incerta, respectively, to form an interface between dorsal and ventral thalami (Fig. 15-7B).
An internal medullary lamina, also consisting of myelinated fibers, extends into the substance of the thalamus, where it forms partitions or boundaries that divide the thalamus into its principal cell groups (Figs. 15-9 and 15-10): the anterior, medial, lateral, and intralaminar nuclear groups. The last cell group is located in the portion of the internal medullary lamina that separates the lateral and medial nuclear groups. In addition, there are midline thalamic nuclei located just superior to the hypothalamic sulcus.
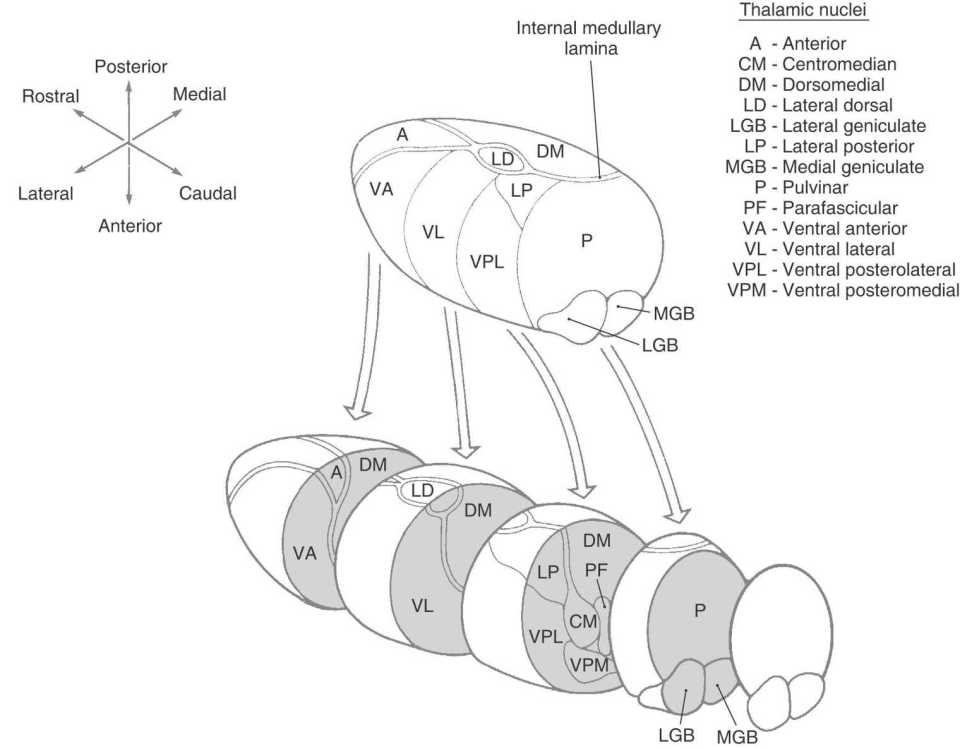
Figure 15-10. Exploded view of the dorsal thalamus illustrating the organization of thalamic nuclei. Compare with Figures 15-6 and 15-8.
Finally, attached to the caudolateral portion of the thalamus are the medial and lateral geniculate bodies (and their correspondingly named subjacent nuclei) (Figs. 15-7D, 15-8D, and 15-10). Although considered here as components of the lateral nuclear group, the geniculate nuclei are sometimes considered as a separate part of the thalamus, the metathalamus.
Anterior Thalamic Nuclei
The anterior nucleus forms a prominent wedge on the rostral aspect of the dorsal thalamus just caudolateral to the interventricular foramen; this wedge is the anterior thalamic tubercle. Internal to the anterior tubercle is a large principal nucleus and two smaller nuclei that collectively form the anterior nucleus of the thalamus (Figs. 15-7A, 15-8B, 15-9, and 15-10). Rostrally, the internal medullary lamina divides to partially encapsulate the anterior nucleus. The cells of this nucleus receive dense limbic-related projections from (1) the mammillary nuclei via the mammillothalamic tract and (2) the medial temporal lobe (hippocampus) via the fornix. The output of this nucleus is primarily directed to the cingulate gyrus through the anterior limb of the internal capsule (Fig. 15-11; see p. 205). The anterior nucleus is an important synaptic station in the Papez circuit, which is related to emotion and memory acquisition.

Figure 15-11. Relationship of the thalamic nuclei with the cerebral cortex as depicted by the patterns of thalamocortical connections. Each thalamic nucleus is pattern coded or color coded to match its target area in the cerebral cortex.
Medial Thalamic Nuclei
This region of the dorsal thalamus comprises the dorsomedial nucleus (Figs. 15-8C and 15-9). This expansive group of neuronal cell bodies is composed of large parvicellular (located caudally) and magnocellular (located rostrally) parts and a small paralaminar part adjacent to the internal medullary lamina (Figs. 15-7B-D and 15-10). The two larger portions are linked to parts of the frontal and temporal lobes and to the amygdaloid complex (Fig. 15-11). Cells of the paralaminar subdivision receive input from the frontal lobe and substantia nigra and may play a role in the control of eye movement.
Lateral Thalamic Nuclei
This large collection of thalamic neurons is grouped into dorsal and ventral tiers. The relatively small group of dorsal tier nuclei includes the lateral dorsal and lateral posterior nuclei along with the much larger pulvinar nucleus (pulvinar) (Figs. 15-7B-D, 15-8D, 15-9, and 15-10). The connections of the lateral dorsal and lateral posterior nuclei are formed with the cingulate gyrus and parietal lobe, respectively (Fig. 15-10). The large pulvinar nucleus consists of anterior, medial, lateral, and inferior subdivisions. The inferior division receives input from the superior colliculus and projects to the visual association cortex. Other portions of the pulvinar project to areas of the temporal, parietal, and frontal lobes that are especially concerned with visual function and eye movements (Fig. 15-11).
The large ventral tier of the lateral group consists of three separate nuclei (Figs. 15-7, 15-8B, C, and 15-9). The ventral anterior nucleus (VA) and the slightly more caudal ventral lateral nucleus (VL) are important motor-related nuclei; the ventral posterior nucleus, consisting of ventral posterolateral (VPL) and ventral posteromedial (VPM) nuclei, convey somatosensory information to the cerebral cortex.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


