FIGURE 32.1 Diagram of the spinal cord and dorsal root showing the peripheral receptors and terminations of fibers within the spinal cord.
The majority of axons originating from second-order spinothalamic neurons cross the midline in the anterior white commissure and gather into the anterior and lateral STs; a small proportion of fibers ascend ipsilaterally. Fibers crossing in the anterior white commissure are affected early in syringomyelia. In the past, anatomists thought the anterior ST carried crude touch and the lateral ST pain and temperature; current evidence suggests all these modalities are carried in both tracts, so the lateral and anterior STs are now sometimes lumped together as the anterolateral or ventrolateral system (ALS) or simply the spinothalamic tract or system. For clinical purposes, it remains useful to consider the pain and temperature pathways in the ST as a distinct system. The ST ascends in an anterolateral position, just medial to the anterior spinocerebellar tract (Figure 32.2). Intermingled with the fibers of the ST are ascending spinoreticulothalamic fibers, which contribute to the ALS. The ST is somatotopically organized, and the distribution of fibers is clinically relevant. Lowermost, sacral and lumbar, fibers entering first are displaced progressively more laterally by subsequently entering fibers. As the tract ascends, the sacral fibers come to lie most lateral and superficial, nearer to the surface of the cord (Figure 32.3), with cervical fibers most medial. There is also a slight rotation so that the sacral fibers also come to lie somewhat more posterior as the tract ascends. At midbrain levels, lower-extremity and sacral fibers are posterior, and those from the upper limb and trunk are more anterior. Since the sacral fibers lie most laterally, an intramedullary spinal cord lesion, such as a neoplasm, may produce “sacral sparing,” preservation of sensation in a saddle distribution in the face of sensory loss otherwise present below a certain spinal level. Conversely, a compressive lesion pressing on the upper spinal cord may preferentially involve the sacral spinothalamic fibers, causing sacral dysfunction first. Fibers carrying deep pain are in general thought to lie nearer the midline than those carrying superficial pain. The spinoreticulothalamic fibers in the ALS subserve diffuse, poorly localized pain from deep and visceral structures. They may also be involved in the affective aspects of pain.
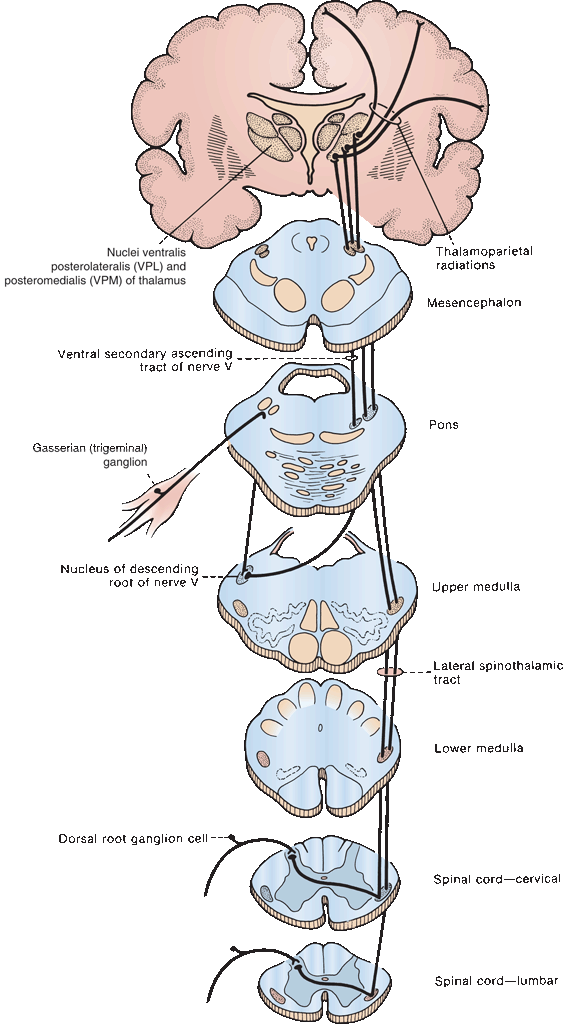
FIGURE 32.2 The lateral spinothalamic tract.
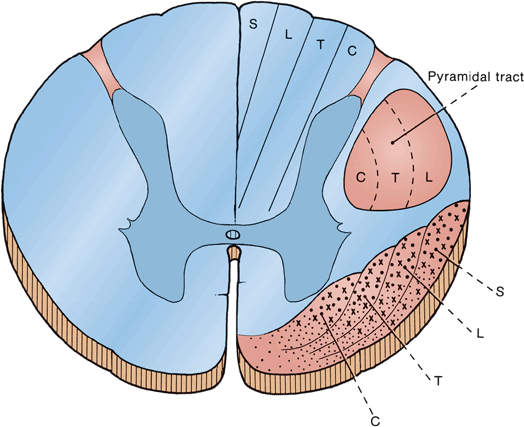
FIGURE 32.3 Diagram of cross section of the cervical region of the spinal cord showing the arrangement of fibers in the spinothalamic and pyramidal tracts and dorsal columns. Heavy dots indicate fibers carrying temperature sensation, crosses indicate fibers carrying pain sensation, and fine dots indicate fibers carrying tactile impulses. C, T, L, and S indicate fibers from or destined for cervical, thoracic, lumbar, and sacral levels of the spinal cord.
In the medulla, the ST lies peripherally, dorsolateral to the inferior olivary nuclei; in the pons, it is lateral to the medial lemniscus (ML) and medial to the middle cerebellar peduncle; in the mesencephalon, it is peripheral, dorsal to the ML and just dorsolateral to the red nucleus. It passes near the colliculi and enters the diencephalon just medial to the brachium of the inferior colliculus.
Pain and temperature fibers from the face enter the pons through the Gasserian ganglion and then descend in the spinal tract of the trigeminal nerve to varying levels, where they synapse on neurons in the adjacent nucleus of the spinal tract (see Chapter 15). These second-order neurons decussate and form the trigeminothalamic tract, which runs near the ascending spinothalamic and lemniscal fibers (Figure 15.2). The other cranial nerves carrying exteroceptive pain sensation have ganglia comparable to the DRG and pathways corresponding to the trigeminothalamic tract. These are discussed in the chapters on the individual cranial nerves.
In the upper lateral midbrain, all the somatosensory fibers begin to converge. The ST fibers are joined in the rostral brainstem by the laterally migrating fibers from the ML and by ascending trigeminothalamic fibers so that ultimately all the fibers subserving somatosensory function run together as they approach the thalamus. The tracts enter the ventrobasal and ventral posterior nucleus of the thalamus together; body sensation fibers terminate in the VPL nucleus and facial sensation fibers in the ventral posterior medial (VPM) nucleus. There is detailed somatotopic organization within VPL and VPM. From the thalamus, fibers run in the thalamic radiations through the posterior limb of the internal capsule to the primary somesthetic cortex in the postcentral gyrus for conscious recognition. The primary somesthetic cortex communicates with the parietal sensory association cortex and with other cortical areas. Thalamocortical fibers also project to the superior bank of the sylvian fissure.
In the thalamoparietal radiations, fibers carrying lower-extremity sensation curve medially to the superior medial surface of the hemisphere adjacent to the medial longitudinal fissure; those from the upper body go to the midportion of the surface of the parietal lobe; those from the face terminate on the lateral, inferior portion of the postcentral gyrus (Figure 6.7). Fibers of the spinoreticulothalamic tract carry nociceptive information in the ALS. There are synapses in the brainstem reticular formation and medial part of the thalamus. Spinoreticulothalamic fibers terminate in the intralaminar thalamic nuclei. The thalamic neurons that mediate pain project both to the parietal lobe and to the limbic cortex. Projections from the intralaminar nuclei terminate in the hypothalamus and limbic system and probably mediate the affective and autonomic responses to pain.
Descending pathways serve to modulate pain. Fibers from the frontal cortex and hypothalamus project to the midbrain periaqueductal gray. The descending pain modulation pathway then descends in the dorsal part of the lateral funiculus to the posterior horn. Descending fibers from the locus ceruleus, the raphe nuclei and other brainstem areas also modulate the pain response. These descending pathways are important in endogenous pain control and opiate analgesia.
Clinical Examination
There are many methods for testing superficial pain sensation. A simple and commonly used method, as reliable as any, is to use a common safety pin bent at right angles so its clasp may serve as a handle. The instrument should be sharp enough to create a mildly painful sensation, but not so sharp as to draw blood. A hypodermic needle is far too sharp unless its point has been well blunted against some hard surface. A broken wooden applicator stick is often used and is usually satisfactory provided the shards are sharp. Adequately sharp ends can be obtained by holding the stick at the very ends while breaking it. Disposable sterile devices, sharp on one end and dull on the other, are commercially available. While it is not necessary for the stimulating instrument to be sterile, whatever is used must be discarded after use on a single patient to avoid the risk of transmitting disease from accidental skin puncture. There is no place in modern neurology for reusable sharp instruments such as the Wartenberg wheel, but disposable pinwheels are available. Various sensory testing devices have been used experimentally. Instruments for evaluating sensation quantitatively are available commercially.
A helpful trick is to hold the pin or shaft of the applicator stick lightly between thumb and fingertip, and let the shaft slide between fingertip and thumb tip with each stimulation. This helps insure more consistent stimulus intensity than putting a fingertip on the end of the instrument and trying to control the force with the hand or wrist. Experience teaches how to gauge the intensity of the applied stimulus and the expected reaction to it. The clinical evaluation of superficial pain, temperature, and touch sensation shows a reasonably good correlation with quantitative assessment.
It is best to do the examination with the patient’s eyes closed. The patient should be asked to judge whether the stimulus feels as sharp on one side as on the other. Always suggest the stimuli should be the same, as by language such as, “Does this feel about the same as that?” Avoid such language as “Does this feel any different?” or “Which feels sharper?” Suggesting there should be a difference encourages some patients to overanalyze and predisposes them to spurious findings and a tedious, often unreliable examination. A commonly used technique is asking the patient to compare one side to the other in monetary or percentage terms, for example, “If this (stimulating the apparently normal side) side is a dollar’s worth (or 100%), how much is this (stimulating the apparently abnormal side) worth?” The overanalytical but neurologically normal patient often responds with an estimate on the order of “95 cents,” while the patient with real, clinically significant sensory loss is more apt to respond with “5 cents” or “25 cents.” Delivering alternately sharp and dull stimuli, as with the sharp and blunt ends of a safety pin and instructing the patient to reply “sharp” or “dull” is frequently useful but may not detect subtle sensory loss only detectable in comparison with an uninvolved area. Slight changes can sometimes be demonstrated in a cooperative patient by asking her to indicate the alterations in sensation when a pinpoint is drawn lightly over the skin. A cooperative patient with a discrete distribution of sensory loss may be able to map out the involved area quite nicely if instructed how to proceed and left alone for a short time with tools and a marking instrument. The affected area can then be compared with a figure showing sensory distributions.
The latent time in the response to stimulation is eliminated and the delineation more accurate if the examination proceeds from areas of lesser sensitivity to those of greater sensitivity rather than the reverse. If there is hypalgesia, move from areas of decreased sensation to those of normal sensation; if there is hyperalgesia, proceed from the normal to the hyperalgesic area. There may be a definite line of demarcation between the areas of normal and abnormal sensation, a gradual change, or at times a zone of hyperesthesia between them. It is occasionally useful to move from the normal to the numb area. In myelopathy, a spinal sensory level that is the same going from rostral to caudal as from caudal to rostral suggests a very focal and destructive lesion; when the two levels are far apart, the lesion is usually less severe. If testing is done too rapidly, the area of sensory change may be misjudged. Applying the stimuli too close together may produce spatial summation; stimulating too rapidly may produce temporal summation. Either of these may lead to spurious findings. If stimulation is too rapid, or if conduction is delayed, a given response may refer to a previous stimulation. Stimuli should be applied at irregular intervals to avoid patient anticipation. If the patient knows when to expect a stimulus, a seemingly normal response can occur even from an anesthetic area. Include control stimuli from time to time, especially if the patient is comparing sharp and dull (e.g., using the dull end of the pin while asking if it is sharp), to be sure the patient has understood the instructions and is paying attention.
Temperature sensation may be tested with test tubes containing warm and cool water, or by using various objects with different thermal conductivity. Ideally, for testing cold, the stimuli should be 5°C to 10°C (41°F to 50°F), and for warmth, 40°C to 45°C (104°F to 113°F). The extremes of free-flowing tap water are usually about 10°C and 40°C. Temperatures much lower or higher than these elicit pain rather than temperature sensations. Normally, it is possible to detect a difference of about 1°C in the range of around 30°C. The tubes must be dry, as dampness may be interpreted as cold. The tines of a tuning fork are naturally cool and work well for giving a quick impression of the ability to appreciate coolness. The tines quickly warm with repeated skin contact; applying the tines alternately and waving the fork in the air between stimuli helps prevent this warming. Holding the tines under cold running tap water may also be helpful. Some examiners warm one tine deliberately by rubbing and then test the ability to discriminate between the warm side and the cool side of the fork. This technique has limited practicality because the cool side warms so rapidly with skin contact. The latency for detecting temperature is longer than for other sensory modalities, and the application of the stimulus may need to be extended.
In the general examination, it is sufficient to determine whether the patient can distinguish hot and cold stimuli. It may be useful in some circumstances, such as the detection of mild peripheral neuropathy, to determine whether the patient is able to differentiate between slight variations in temperature. This is best done with special devices for testing temperature sensation quantitatively. In most instances, heat and cold sensibility are equally impaired. Rarely, one modality may be involved more than the other; the area of impaired heat sensibility is usually the larger. Pain and temperature sensibility are usually involved equally with lesions of the sensory system, and it is rarely necessary to test both. Testing temperature may be useful when the patient does not tolerate pinprick stimuli, has confusing or inconsistent responses to pain testing, or to help map an area of sensory loss. In some instances, the deficit is more consistent with temperature testing than with pinprick. Temperature testing may not be very reliable in patients with circulatory insufficiency or vasoconstriction causing acral coolness.
Quantitative sensory testing (QST) uses neuro-physiologic methods to examine sensation. It provides very accurately measured stimuli of various types and uses strict paradigms for recording responses. Temperature sensation is tested by delivering pulses of hot and cold and determining the threshold for detection. Extremes of temperature assess pain. There is good correlation between QST and clinical methods, but QST is very useful for longitudinal studies.
TACTILE SENSATION
Anatomy and Physiology
Cutaneous receptors that mediate light touch or general tactile sensibility include free nerve endings, Merkel cell endings, and encapsulated endings such as Meissner’s and Pacinian corpuscles and Ruffini endings. All the encapsulated receptors function as mechanoreceptors with afferent nerve fibers in the group II and III range. Pacinian corpuscles are large, lamellated structures located subcutaneously in the palmar, plantar and digital skin, genitalia, and other sensitive areas; they function as rapidly adapting mechanoreceptors. They are especially responsive to vibration, most notably in the 40 to 1,000 Hz frequency range. Meissner tactile corpuscles are found primarily in thick hairless skin, such as the hand, foot and lips, and are most highly developed in the finger pads. They also respond to vibration in the low-frequency range (10 to 400 Hz) and are maximally sensitive at 100 to 200 Hz. Merkel cell receptors are also slowly adapting mechanoreceptors that respond to low frequency vibration. Ruffini endings are slowly adapting mechanoreceptors located in hairy as well as glabrous skin, in joint capsules, tendon insertions, and elsewhere. They are particularly responsive to stretching or indentation of the skin.
Light touch sensation is conveyed over large and small myelinated peripheral nerve fibers to unipolar DRG cells. The neurons subserving fine discriminative touch are the largest cells in the DRG. Tactile sensation follows several different pathways within the central nervous system. The central processes enter the spinal cord via the medial division of the posterior roots, and bifurcate into ascending and descending fibers (Figure 32.4). Fibers carrying fine discriminatory and localized tactile sensibility then, without synapsing, turn upward in the ipsilateral posterior column. Fibers carrying crude touch synapse within several segments of their point of entry, and the axons of the neurons of the next order cross to the opposite ALS. Other tactile fibers have a synapse in the posterior horn, and ascend in the dorsolateral funiculus to the lateral cervical nucleus at C1-C2, where axons of the next order neurons decussate and join the ML. In the posterior columns, fibers from the lumbosacral region aggregate near the midline, and fibers from successively more rostral regions aggregate in a progressively more lateral position, producing somatotopic lamination, the reverse of the STs (Figure 32.3). In the STs, the sacral fibers are most lateral; in the posterior columns, the lowest fibers are most medial. All the fibers below about T8 are grouped together in the fasciculus gracilis; analogous fibers above T8 form the fasciculus cuneatus.
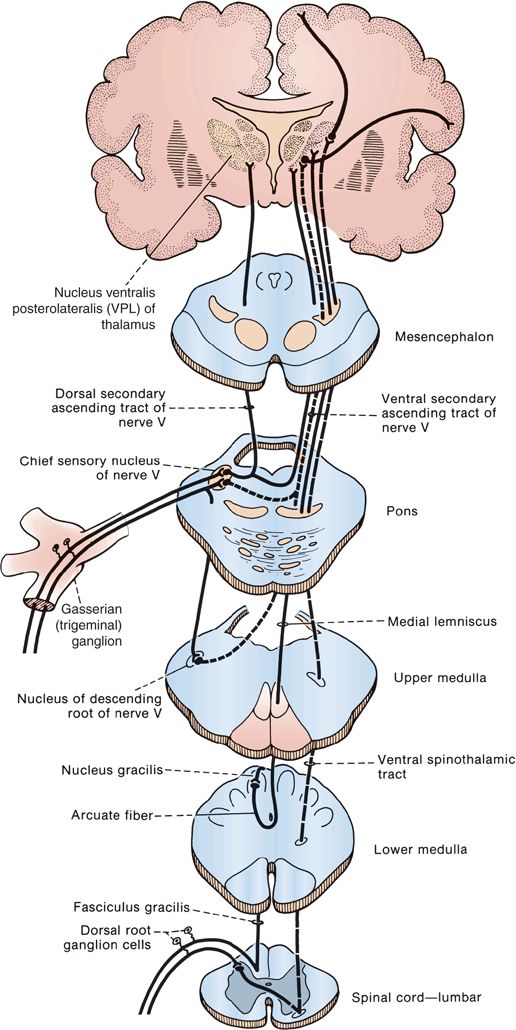
FIGURE 32.4 The tactile pathways.








