Fig. 11.1
The monosynaptic stretch reflex. (a) In response to rapid stretch sensory Ia afferents activate alpha motoneurons in the ventral horns of the spinal cord resulting in a delayed contraction of the muscle that was stretched. Clinicians make use of this response to test spinal cord reflexes. (b) The same pathways can be assessed intraoperatively using the H-reflex. A peripheral nerve is electrically stimulated, producing two responses that can be recorded with EMG. An early response, known as the M-wave, is elicited by direct activation of the muscle via motor axons. A later response, the H-reflex, is the result of activation of sensory Ia afferents, similar to what occurs when the muscle is stretched
Electrically Evoked Responses
Unlike the stretch reflex that is detected by visual observation, EMG is used to record the M-wave, H-reflex, and F-response. In EMG testing, muscle contractions are recorded as compound muscle action potentials (CMAPs). Electrophysiological recordings afford the clinical scientist the opportunity to make precise measurements of latency, amplitude, and morphology (Fig. 11.1b).
Two physiological differences distinguish the H-reflex from the stretch reflex: (1) the H-reflex is evoked by electrical stimulation of a mixed motor and sensory nerve rather than by muscular stretch and (2) the H-reflex is activated proximal to the muscle and avoids entirely the muscle spindle fibers which, along with gamma motor neurons, play a role in modulating stretch-reflex gain. These factors make the H-reflex well suited to assessing spinal cord excitability [7].
The H-Reflex
Electrical activation of a mixed peripheral nerve creates an action potential that propagates in both directions along both sensory and motor axons (i.e., both ortho- and antidromically in afferent and efferent axons). The stimulation threshold for the H-reflex is typically low, and the reflex response is characterized by consistent latency between trials and simple morphology, leading to the conclusion that the reflex is mediated by large-diameter, monosynaptic Ia afferent fibers [5]. Despite this, there is some evidence for oligosynaptic components to the H-reflex response [6].
In humans, the CMAP response evoked by the lowest intensity stimulation is likely to be the H-reflex. The stimulus intensity where the H-reflex is first recorded is near or below the motor threshold, and therefore, an orthodromic motor response (M-wave) may not be recorded.
The H-reflex response is most like the muscle stretch reflex as it is evoked by the same process whereby a signal travels orthodromically along Ia sensory afferents towards the spinal cord, crosses the synapse onto alpha motor neurons, and then travels orthodromically along efferent motor axons to the muscle where it evokes a delayed muscular contraction [5]. Because of this similar route, it shares a similar characteristic delay with the stretch reflex. H-reflexes evoked at the popliteal fossa and recorded at the soleus muscle typically have a latency of ~30 ms, while those evoked at the cubital fossa and recorded at the flexor carpi radialis muscle have a latency of ~18 ms [8]. As with the stretch response, this delay is due to the longer route that this signal must take.
The M-Wave
As stimulation intensity is gradually increased, a shorter latency CMAP begins to appear in the recording. This response is termed the M-wave and is activated not through a reflex circuit but via the direct orthodromic transmission of an action potential along the motor axon to the neuromuscular junction. The stimulus intensity where the M-wave is first recorded is the termed the motor threshold. The M-wave response has the shortest latency because it is the simplest physiologically, being the result of the direct activation of the motor axon and subsequent transmission of an action potential to the neuromuscular junction, producing a contraction of the postsynaptic muscle. As stimulus intensity increases further, the H-reflex will peak in amplitude and then begin to decline as the M-wave increases. Near supramaximal stimulation intensities, the M-wave dominates the recording as its amplitude peaks and the H-reflex disappears altogether.
The F-Response
By the time stimulus intensity becomes supramaximal, a third CMAP response appears on the EMG recording with a similar latency to the H-reflex. Termed the F-response, this response is not a reflex but is generated by an action potential that travels first antidromically and then orthodromically along motor axons. As just explained, the initial orthodromic action potential generated by electrical stimulation will generate a short-latency response, the M-wave. However, the same motor axons will also generate antidromic action potentials that travel towards the spinal cord along the same axons. When the antidromic action potential reaches the motor neuronal pools, the majority of these action potentials will be abolished. However, some of these signals will survive to depolarize the cell body causing an orthodromic action potential to form and travel back down the same motor axons. This “backfiring” of the motor neuron results in a CMAP response in the EMG recording. The population of motor units recruited to produce an F-response will vary from trial to trial yielding variable amplitude, latency, and morphology. This is one way in which the F-response can be distinguished from the H-reflex [9].
Ordered Responses Explained
The H-reflex, M-wave, and F-response are recruited in an ordered manner by electrical stimulation of increasing intensity. This occurs because the excitability of axons when evoked by electrical current is directly related to their diameter and input resistance; the largest axons will be recruited by the lowest stimulus intensity [10]. The largest diameter axons in a mixed peripheral nerve are the Ia afferent axons responsible for carrying the sensory action potential which initiates the H-reflex. The second largest group of axons are those of the alpha motor neurons, especially those that innervate larger, fast-twitch fatigable motor units in skeletal muscle. There is some overlap in the diameters of these axons, which explains why there is also some overlap in the intensities at which the M-wave and H-reflex are recorded. Nevertheless, the H-reflex is typically first noted at stimulus intensities that are subthreshold for the M-wave.
Advantages of the H-Reflex
Because H-reflexes are single-sweep and do not require averaging they offer a real-time test, similar in this respect to TcMEPs. They are also like TcMEPs in that they involve spinal cord circuitry in the gray matter; however, unlike TcMEPs they can be run without having to pause or interrupt the surgery as they produce little or no detectable movement. Furthermore, they have been shown to be stable with anesthetic regimens commonly employed to allow intraoperative monitoring [11].
Perhaps the greatest physiological advantage of H-reflexes is that they can be used to assess not just the nerve roots through which the afferent and efferent signal travels but complex suprasegmental, propriospinal, and interneuronal circuitry that affects the reflex arc both pre- and postsynaptically [6]. When evoked by stimulation of the posterior tibial nerve at the popliteal fossa, or the median nerve at the cubital fossa, H-reflexes can be minimally understood to be providing information about the integrity of S1 and C6/C7 nerves and nerve roots, respectively. However, the potential advantage of H-reflex monitoring is that it may provide a way of monitoring the integrity of a much larger network of suprasegmental spinal cord circuitry. Leppanen has speculated that the loss of H-reflexes following spinal cord trauma may have to do with uncoupling of the central pattern generator in humans and the disruption of inputs onto segmental afferents, yielding a change in reflex gain [8]. Although this is an intriguing hypothesis, it is difficult to be certain about the specific mechanisms of H-reflex suppression in humans following spinal cord trauma.
Two reports using H-reflexes in the operating room have described H-reflexes as being remarkably sensitive to intraoperative events. Standard surgical maneuvers such as hammering with a mallet, distraction, and derotation of the spine resulted in transient decreases in H-reflex amplitude [12, 13]. This decrease in H-reflex amplitude was repeatedly observed across multiple procedures and was correlated with stressful spinal manipulations and perturbances of the spinal cord. The authors of this chapter have observed similar decreases in H-reflex amplitude correlated with spinal corrections or EMG bursts observed during posterior decompressions (Fig. 11.2).
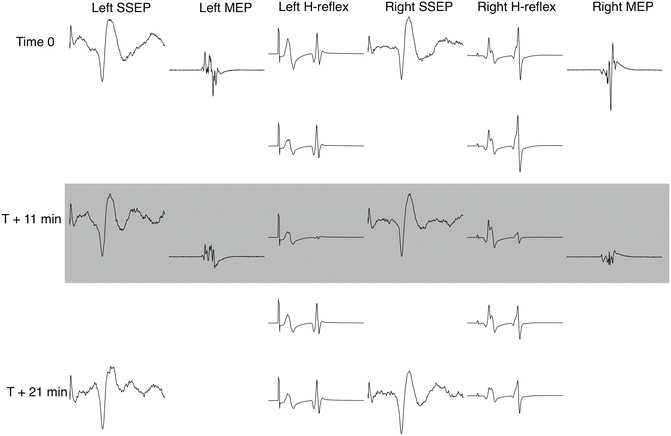
Fig. 11.2
The H-reflex is sensitive to spinal irritation. Displayed signals were gleaned during a complex scoliosis correction in an 18-year-old male. SSEP and H-reflex tests were gathered at regular intervals, while the TcMEP was run as often as practical, in communication with the surgical team. Pictured signals include bilateral cortical SSEPs evoked from the posterior tibial nerve, bilateral H-reflexes recorded at the soleus muscle, and bilateral TcMEPs recorded at the abductor hallucis muscle. During osteotomy, a large EMG burst (not shown) was observed across multiple lower limb muscles bilaterally in response to a distinct hammer strike upon the osteotome. The surgical team noted the same response as a brief but large patient spasm and requested TcMEPs to be tested. H-reflexes were significantly diminished for a period of approximately 5 min bilaterally. SSEPs remained undiminished while TcMEPs were diminished in amplitude but remained present in all recorded muscles bilaterally. Both H-reflexes and TcMEPs were determined to be unchanged from baselines at close and the patient awoke with no deficit
Practical Considerations
Anesthesia
As with other intraoperative modalities, H-reflex and F-response data can be compromised by anesthetic regimens that are not optimized to provide the best environment for achieving valid neurophysiological results. Critically, H-reflexes and F-responses rely upon accurate recordings of muscular contraction via EMG. As such, they are strongly affected by paralytics applied during surgery. Neuromuscular blocking agents will diminish or even abolish the CMAP responses evoked as H-reflexes or F-responses. Interpretation of H-reflexes and F-responses should take into account the degree of neuromuscular blockade.
Related posts:
 New Vistas: Small-Pain-Fibers Method of Testing for Spinal Cord Assessment in Pain States
Neurological Assessment and Correlation in Spinal Cord Nerve Root Pathology
New Vistas: Small-Pain-Fibers Method of Testing for Spinal Cord Assessment in Pain States
Neurological Assessment and Correlation in Spinal Cord Nerve Root Pathology
 Electromyography (EMG)
Electromyography (EMG)
 Intraoperative Monitoring for Carotid Endarterectomy
Intraoperative Monitoring for Carotid Endarterectomy
 Peripheral Nerve Monitoring
Peripheral Nerve Monitoring
 Spinal Cord Stimulation: Principles and Applications
Spinal Cord Stimulation: Principles and Applications
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


