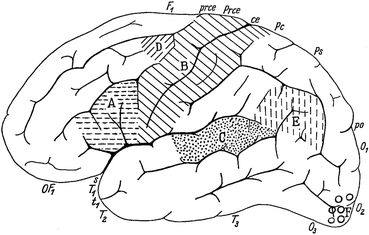
Fig. 3.2
Lateral view of the human telencephalon. Areas, the functions of which were known at the end of the nineteenth century are marked. A, motor speech centre of Broca; B, the somatomotor and somatosensory areas; C, auditory area; D, center concerned with writing; E, Wernicke’s sensory speech centre; F, visual area (Modified from Vogt and Vogt 1954)
The approach chosen by the Vogts was relatively simple and remained essentially unchanged throughout their long scientific career (Vogt 1903, 1943; Vogt and Vogt 1919, 1936, 1942, 1954, 1956). It included a systematic analysis of those structural features of the cerebral cortex that can be readily recognized with relatively weak magnifications, with a view at identifying and delimiting fundamental morphological units within that organ, assuming that the units or areas of distinct structure thus identified, would also proven to be organs of special function. The results of these architectonic studies were expected to provide an adequate basis for clinicopathological studies, as well as for the study of the brains of geniuses (‘Elitegehirne’), and feeble-minded people.
At the time that the Vogts began their studies there were two staining procedures for brain tissue, which yielded reliable and reproducible results, i. e. the Nissl stain for neuronal cell bodies, and the Weigert stain and its variants for myelinated nerve fibres. Thus, it became routine in the Neurologisches Zentralstation to prepare serial sections of human and animal brains, and to stain these series according to the two procedures mentioned. The systematic study of the material thus prepared led to the emergence of two new neuroanatomical subdisciplines, which were designated by O. Vogt (1903) as cytoarchitectonics and myeloarchitectonics.
The cytoarchitectonics of the cerebral cortex became the specialism of Korbinian Brodmann, who joined the Vogts in 1901 and remained attached to their laboratory until 1909. Brodmann studied the cellular structure of the cortex in a considerable number of mammals, including the hedgehog, the flying fox, the lemur, the guenon, and the human, resulting in an impressive series of publications (Brodmann 1903a, b, 1905a, b, 1906, 1908a, b), and in a summarizing monograph (Brodmann 1909). His famous map of the cytoarchitecture of the human cerebral cortex was first published in Brodmann (1908a), and its final version appeared in Brodmann (1914). It is of note that the practising of the cytoarchitectonics of the human cerebral cortex remained by no means confined to the institute of the Vogts. Von Economo and Koskinas (1925, Vienna), Bailey and Von Bonin (1951, Urbana, Illinois) and Sarkissov et al. (1955, Moscow) all published comprehensive works on the human cortex, resulting in complete cytoarchitectonic parcellations of that organ (see Braak 1980 and Zilles and Amunts 2010, for synopsis and further developments).
Study of the myeloarchitectonics of the cerebral cortex, on the other hand, remained largely concentrated in the laboratories of the Vogts, from its beginning, marked by the appearance of O. Vogt’s (1910a) preliminary note on the human frontal cortex, to its end, marked by the appearance of Hopf’s (1970b) study of the human parietal cortex. The comprehensive program included analyses of the myeloarchitecture of all parts of the human cortex, as well as the cortices of a number of mammals, including the hedgehog (Flores 1911), the mangabey (Mauss 1908), the orang-utan (Mauss 1911) and the chimpanzee (Beck 1929; Strasburger 1937b; Gerhardt 1938).
3.2 The Principles of Myeloarchitectonics
Preparations stained with the Weigert method reveal that the neocortex contains numerous myelinated fibres, which show two principal orientations, tangential and radial. The tangential fibres tend to form local concentrations or bands, the most conspicuous of which can be clearly observed with the naked eye in unstained sections. The radial fibres are concentrated in bundles or radii. O. Vogt (1903) designed a basic plan (‘Grundschema’) of the myeloarchitectonic organization of the neocortex, which formed the point of departure of all myeloarchitectonic studies produced in his laboratory. This scheme is shown in Fig. 3.3, together with a comparable basic scheme of the cytoarchitectonic organization. In both schemes the neocortex is subdivided into six layers. The cytoarchitectonic layers are: (I) the cell-poor zonal layer, (II) the external granular layer, (III) the external pyramidal layer, (IV) the internal granular layer, (V) the internal pyramidal layer, and (VI) the multiform layer. The corresponding myeloarchitectonic layers are, in order to avoid confusion, designated with Arabic, rather than with Roman numerals.
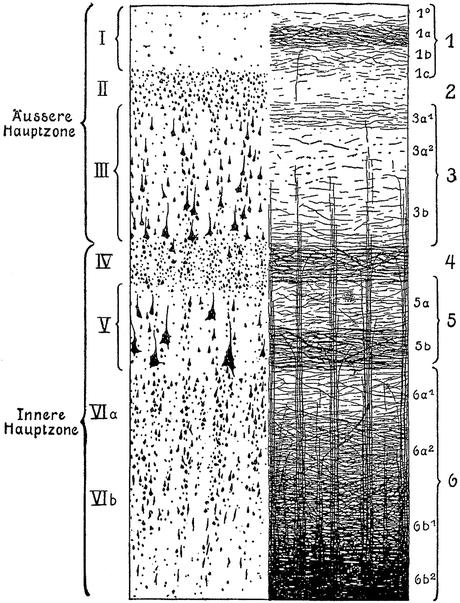
Fig. 3.3
O. Vogt’s (1903) basic schemes of the cytoarchitectonic layers (designated with Roman numbers), and the myeloarchitectonic layers (designated with Arabic numbers)
1.
The zonal layer is differentiated into four sublayers, the narrow sublayer 1°, which contains only very few fibres, and the external, intermediate and deep sublayers 1a, 1b and 1c, of which 1a contains clearly more fibres than 1b and 1c.
2.
The dysfibrous layer which contains, just like sublayer 1°, only very few fibres.
3.
The suprastriate layer has three sublayers, of which the superficial sublayer 3a1 is more rich in fibres than the remaining sublayers 3a2 and 3b. In certain cortical regions, sublayer 3a1 show a distinct concentration of fibres, known as the stripe of Kaes-Bechterew. Sublayer 3b is characterized by the presence of the end-segments of the radial bundles.
4.
The external stria or outer stripe of Baillarger forms a dark band of tightly packed, tangential fibres.
5a.
The intrastriate layer is generally relatively poor in tangential fibres, thus contrasting with the bordering stripes of Baillarger.
5b.
The internal stria or inner stripe of Baillarger is again a dense plexus of tightly packed tangentially oriented fibres.
6.
This layer is subdivided into the pale substriate lamina 6a1 and laminae 6a2, 6b1 and 6b2, which show an increasing wealth of tangentially oriented fibres. Sublayer 6b2 forms the zone of transition to the subcortical white matter.
Variations in the number and density of the tangential and radial fibres define the boundaries of te myeloarchitectonic areas. With regard to the lamination of the tangential fibres, O. Vogt (1910a, b, 1911) distinguished four principal types (Fig. 3.4).
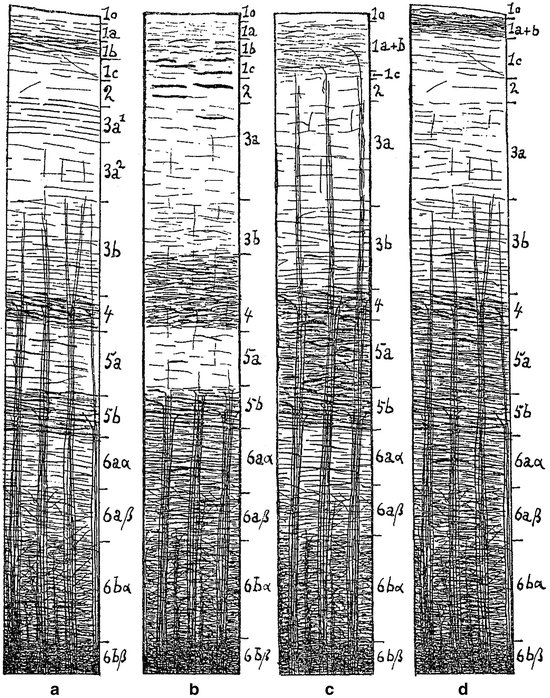
Fig. 3.4
The four principal types of myeloarchitectonic layering, according to O. Vogt (1910a). (a), bistrate type; (b), unistriate type; (c), unitostriate type; (d), astriate type
(a)
A bistriate type, characterized by the presence of two distinct and clearly separated bands of Baillarger (Fig. 3.4a). It should be added that the situation in which the density of fibres in the outer stripe equals that in the inner stripe is designated as typus equodensus, and that the situations in which the fibres in the outer stripe are more densely or less densely arranged than those in the inner stripe, are designated as typus externodensior and internodensior, respectively.
(b)
A unistriate type, in which only the external stripe of Baillarger can be distinguished as a separate entity. The inner stripe, though present, cannot be delineated because of the high fibre content of the substriate lamina 6a1 (Fig. 3.4b, in which the substriate lamina is labeled 6aα).
(c)
A unitostriate type, in which the fibre-poor interstriate layer is lacking and the two stripes of Baillarger form a single plexus (Fig. 3.4c).
(d)
An astriate type, in which, due to the presence of an unusually large number of tangential fibres in intrastriate layer 5a and substriate lamina 6a1, layers 4–6 form a single dark and undivided fibre zone (Fig. 3.4d).
As regard the disposition of the bundles of radial fibre bundles, O. Vogt distinguished three types, euradiate, supraradiate and infraradiate. In the euradiate type, the radii do not extend beyond the level of the suprastriate layer (Fig. 3.4a, d); in the supraradiate type, the bundles traverse almost the entire width of the cortex and reach the zonal layer (Fig. 3.4c), whereas in the infraradiate type, the radii are very short and terminate already in the fifth layer at the level of the inner stripe of Baillarger (Fig. 3.4b).
The radii do not vary only in length, but also in breadth, number and calibre of their fibres. On the basis of differences in the breadth of radii, latoradiate, medioradiate and tenuiradiate types were distinguished. Differences in the number of radii led to distinction of densoradiate, modicoradiate and sparsoradiate types, whereas differences in the size of the fibres forming the radii found their expression in grossoradiate and finoradiate types.
O. Vogt codified many other myeloarchitectonic variations as types. Thus, he referred to an overall wealth of myelinated fibres in a given area as typus dives, whereas an overall scarcity of such fibres was characterized as typus pauper. The various tangential layers generally consist of a plexus of thin fibres of about equal size (‘Grundfasern’), in which individual fibres of larger size (‘Einzelfasern’) are embedded. O. Vogt designated layers, in which the coarse individual fibres are scarce, as representing a typus tenuifibrosus, and layers in which these fibres are numerous as representing a typus grossofibrosus.
It should be appreciated that the myeloarchitectonics of the cortex, as developed by O. Vogt, in spite of its extremely detailed and intricate typology, remained a purely descriptive and qualitative neuroanatomical subdiscipline. Several attempts at the development of a more quantitative and more objective myeloarchitectonics have been made. Thus, Braitenberg (1962) devised a method in which the light absorption, being directly proportional to the fibre density, is systematically measured in narrow strips of Weigert-stained sections of the cerebral cortex, passing from the pial surface to the white matter. With this method, he recorded the fibre density in sections taken from 14 different cortical regions. Representation of the results in graphic form yielded quite characteristic curves for most of the regions studied (Fig. 3.5a, b). Braitenberg also produced an interesting diagram, explaining the relationships between the basic functional wiring and the myeloarchitecture of the cortex, and the overall course of his photometric curves (Fig. 3.5c). Hopf (1966) developed a photometric method for determining the extent of myelinization in the cortex, which closely resembled that of Braitenberg. With the aid of this technique, he successfully explored the myeloarchitecture of the human frontal, temporal and parietal cortices (Hopf 1968a, b, 1969, 1970b).
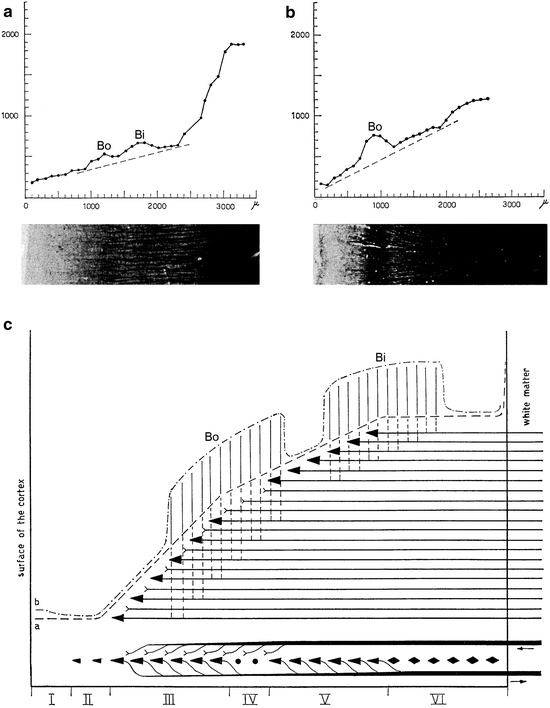
Fig. 3.5
Braitenberg’s (1962) photometric analysis of the myeloarchitecture of the human cerebral cortex. (a, b) Graphic representations of photometric analyses of strips of myelin-stained sections of the cortex, with distance from the surface of the cortex (in mμ) on the abscissa, and fibre-density (in arbitrary units) on the ordinate. The tangents of the curves show different slopes. The outer and inner stripes of Baillarger (Bo, Bi) produce local prominences over the tangents. (a) Cortex of middle frontal convolution, showing the presence of both stripes. (b) Area striata with the pronounced outer stripe of Baillarger (or line of Gennari, or line of Vicq d’Azyr), after which it is named. (c) Diagram, clarifying the relationships between the overall course of the photometric curves, the myeloarchitecture, and the basic functional wiring of the neocortex. The following hodological features are taken into account: (1) The number of specific afferent fibres in the cortex decreases if we ascend from the white matter; most of these fibres form their terminal ramifications in layers III and IV. (2) The number of efferent fibres increases if we descend to the white matter; practically all of these fibres are produced by pyramidal neurons, situated principally in layers III and V. These two features explain the steady increase in myelinization if we pass in the cortex from superficial to deep, and therewith the overall course of curve a. (3) The axons of the pyramidal neurons produce long, tangentially running collaterals, which tend to assemble at two different cortical levels. It is these concentrations of pyramidal collaterals that, forming the stripes of Baillarger, produce the local increases in fibre density shown in curve (b)
In what follows, the literature on the myeloarchitecture of the human cerebral cortex, as produced by the Vogts and their numerous disciples, will be reviewed first. Next, a general concept, concerning the organization of the cerebral cortex, which has been developed from the myelarchitectonic studies reviewed, will be discussed, and finally, some remarks will be made on the functional parcellation of neocortex. It is felt appropriate, however, to preface this long story with a few remarks on the work of three myeloarchitectonic pioneers, who did not belong to the Vogt-Vogt-school.
3.3 Notes on the Work of Three Myeloarchitectonic Pioneers
3.3.1 Alfred Walter Campbell (1868–1937)
The pathologist Campbell published in 1905 a monograph, entitled: “Histological studies on the localisation of cerebral functions”. The opening paragraph, which gives a clear view on the perspective in which the author placed his work, may be quoted in full: “The process leading to the accomplishment of functional localisation in the cerebral cortex is such a complicated one, and involves so many side issues, that perfection cannot be attained or even hoped for until the fruits of investigation in a number of departments are thoroughly weighed, sifted and assorted. It is anticipated that the observations set forth in this research will help to establish the value of histological work as an auxiliary force in the final settlement of that functional subdivision of the cerebral cortex at which we aim” (l.c. p. XV).
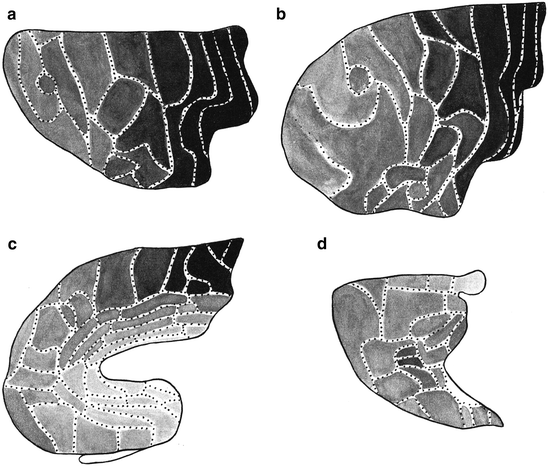
The normal human material, on which the work is based, consisted of three cerebral hemispheres completely examined for both nerve cells and nerve fibres, and three additional hemispheres examined for fibres only. Campbell distinguished 16 cortical areas, which he designated either with topographical (such as: frontal, postcentral, and temporal), or with (provisional) functional names (such as: olfactory, visuo-sensory and visuo-psychic). All of these areas were described in some detail, and their cytoarchitecture and myeloarchitecture were recorded in beautiful drawings (Fig. 3.6c, d). The resultant map, which represents the only complete, combined cyto- and myeloarchitectonic map of the human cerebral cortex, produced thus far, is shown in Fig. 3.6a, b. It will be seen that in Campbell’s parcellation, the frontal cortex is unusually large; that the intermediate-central cortex, which roughly corresponds to Brodmann’s area 6, is not demarcated from Broca’s motor speech region (areas 44 and 45 of Brodmann), and that the temporal area does not only occupy the inferior and middle temporal convolutions, but rather extends over vast regions of the parietal lobe.
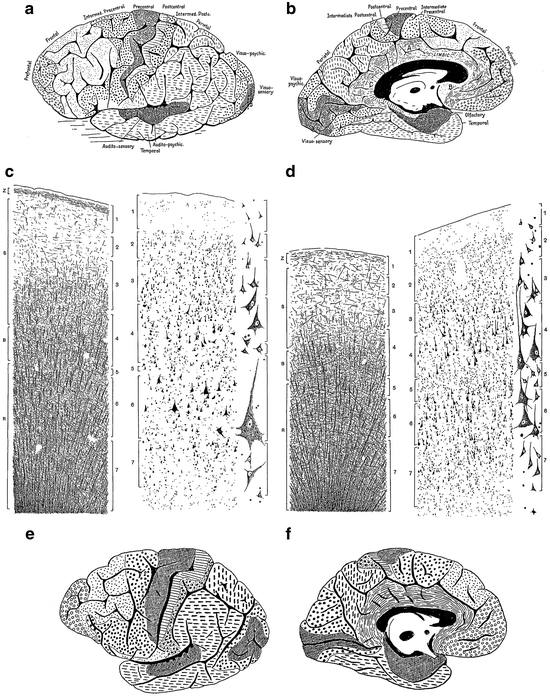
Fig. 3.6
Some illustrations from Campbell’s (1905) monograph: “Histological Studies on the Localisation of Cerebral Function”. (a, b) Architectonic map of the human cerebral cortex; (a), lateral view; (b), medial view. (c, d) Drawings showing the structure of the precentral or motor area (c), and the postcentral or somatosensory area of the human cerebral cortex (d). In each of these figures the pattern of myelinated fibres is shown to the left, and the arrangement of the neuronal cell bodies to the right. Representative cell types are shown at a higher magnification to the far right. (e, f) Architectonic map of the cerebral cortex of the chimpanzee
Campbell did not confine himself to the human brain, but also studied and mapped the cortex of several mammals, including the cat, the orang-utan and the chimpanzee (Fig. 3.6e, f).
3.3.2 Grafton Elliot Smith (1871–1936)
The anatomist Elliot Smith published in 1907 a detailed map of the human cerebral cortex, which, surprisingly, was exclusively based on macroscopic observations (Fig. 3.7a, b). He made fresh sections at many locations of the hemisphere, using differences in the width and distinctness of the stripes of Baillarger as the main criteria for his parcellation. In unstained preparations, these stripes can be recognized as whitish bands, contrasting with the darker hue of the cortical grey matter (Fig. 3.7c). With the aid of this simple technique, Elliot Smith was able to distinguish about 50 different cortical areas. He noticed that most of these areas have precise relations to various stable sulci. As regards the nature of the interareal boundaries, Elliot Smith (1907, p. 240) took a firm stand: “There is a very widespread belief that the characters of an area merge gradually and imperceptably into those of the neighboring areas, but this is entirely mistaken. The changes in structure occur with the utmost abruptness, so that it is possible to determine with absolute precision the exact boundaries of each area,”
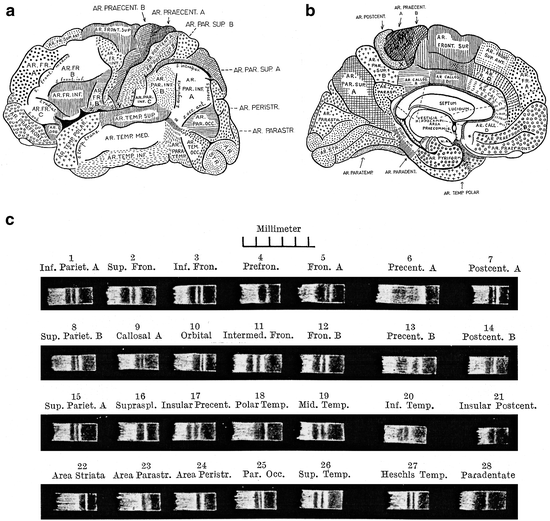
Fig. 3.7
Elliot Smith’s (1907) ‘myeloarchitectonic’ analysis of the human cerebral cortex. (a, b) Architectonic map, based entirely on the study of fresh, unstained macroscopic sections of the cerebral cortex. (c) Pictures showing 28 of the about 50 areas, distinguished by Elliot Smith
3.3.3 Theodor Kaes (1852–1913)
Kaes was psychiatrist and prosector at the asylum Friedrichsburg near Hamburg. Between 1891 and 1904, he published a series of papers on the various techniques, used for the staining of myelinated fibres, and on the myeloarchitecture of the human cerebral cortex. He summarized his findings and views in his opus magnum (Kaes 1907), entitled: “Die Grosshirnrinde des Menschen in ihren Massen und ihrem Fasergehalt. Ein gehirnanatomischer Atlas mit erläuterndem Text”, comprising, apart from a concise text, an atlas consisting of 90 large colour plates, showing the myeloarchitecture of 12 selected cortical regions in 45 individuals, and numerous tables and curves, documenting thousands of measurements of the total width of the cortex, and of its various layers and zones.
Kaes collected 45 human brains, ranging from three months postnatal to 97 years, among which several mentally retarded and criminal individuals. He selected 12 regions in both hemispheres of these brains for further analysis. These regions were designated as: (1) vordere Stirne, (2) hintere Stirne, (3) vordere Zentralwindung, (4) hintere Zentralwindung, (5) Operculum, (6) Insel, (7) vordere Schläfe, (8) hintere Schläfe, (9) oberer Scheitel, (10) unterer Scheitel, (11) Sehrinde, and (12) Gyrus fornicatus. Most unfortunately, any further indication concerning the exact location of the regions selected, is lacking in the work. Sections, taken from these 12 regions, were stained according to the Weigert-Wolters technique. The best-stained sections were carefully drawn and included in the atlas. Thus, in the atlas 24 sections of the cortex of each individual investigated were included, 12 from the left, and 12 from the right hemisphere (Fig. 3.8).
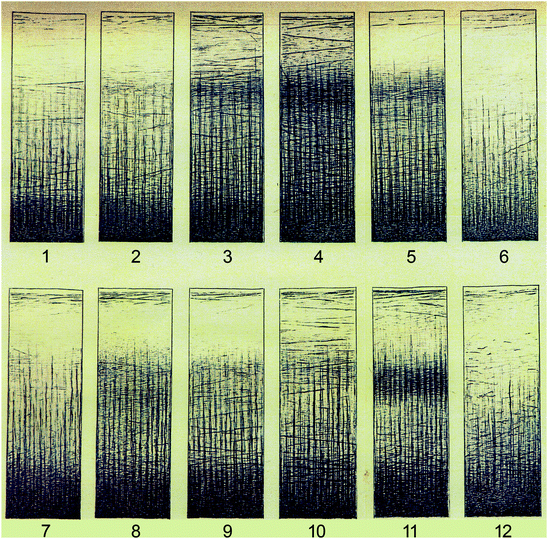
Fig. 3.8
One of the 90 coloured plates, with which Kaes (1907) illustrated his great work on the human cerebral cortex, reproduced at half of the original size. The plate shows the structure of the cortex, as seen in preparations stained with Wolters’ variant of the Weigert technique, in 12 representative cortical areas. The numbers are specified in the text
Kaes used the material, thus selected, for a qualitative and quantitative analysis of the postnatal development of the cortex under normal and abnormal circumstances. In his quantitative studies, he divided the cortex into an external principal zone (‘äußere Hauptzone’), encompassing layers I-III, and an internal principal zone (‘innere Hauptzone’), consisting of layers IV-VI (Fig. 3.3). A detailed discussion of the results of Kaes, falls outside the scope of the present review. Hence, I confine myself to some of his main conclusions: (1) Cortices, which are poor in myelinated fibres, are in general wider than cortices containing numerous fibres. (2) The internal principal zone attains the peak of its development around the 19th year of life; the external principal layer continues developing until the 45th year of life and beyond. (3) The findings on brains of mentally retarded individuals (‘Idiotengehirne’), confirm the rule, mentioned above under 1. (4) The brains of criminals show generally an abnormally low weight, and an infantile level of development.
3.4 Myeloarchitectonic Parcellations of the Human Neocortex
3.4.1 The Frontal Lobe
The literature on the myeloarchitecture of the frontal lobes is voluminous and encompasses studies of O. Vogt (1910a, b), C. and O. Vogt (1919), Strasburger (1937a, b, 1938), Braitenberg (1956), Hopf (1956, 1968a), and Sanides (1962, 1964). The myeloarchitectonic parcellation of this lobe, presented by O. Vogt (1910a), is complex (Fig. 3.9). He distinguished six regions, which were designated with Roman numerals. Each of these regions was subdivided into several (two to four) subregions, and these were, in their turn, further subdivided into divisions, and locally even still further into subdivisions. Finally, one or several areas were delineated within each of the (sub) divisions. In total, 66 myeloarchitectonic areas, designated with Arabic numerals, were distinguished within the frontal lobe. These numerals have nothing to do with the – also Arabic – numerals, used by Brodmann (1909) for his cytoarchitectonic areas. Each of the entities distinguished was designated with a full Latin name, referring to particular myeloarchitectural features characterizing that particular entity. A survey of O. Vogt’s nomenclature for region III and its subdivisions is presented in Table 3.1. O. Vogt does not specify the histological material he used in this study. Strasburger (1937b) mentioned that it was principally based on serial sections of a single hemisphere, designated as A 18r. Vogt’s (1910a) paper ends abruptly after the description of the last area (Fig. 3.10).

III. Regio unistriata euradiata grossofibrosa (areae 33–43) |
|---|
α Subregio unistriata progrediens |
αα Divisio pauper |
33 area latofasciculata |
34 area sublatofasciculata |
35 area aequofasciculata |
αβ Divisio dives |
36 area subtenuistriata |
37 area aequostriata |
β Subregio propeastriata |
38 area subunistriata |
39 area propeastriata |
γ Subregio unistrata degrediens |
40 area dives |
41 area pauper |
δ Subregio astriata |
42 area typica |
43 area atypica |
Fig. 3.10
The end of O. Vogt’s (1910a) preliminary study on the myeloarchitectonic parcellation of the human frontal lobe
In a subsequent publication, O. Vogt (1910b) comments briefly on the findings just reviewed, in relation to the results of the cytoarchitectonic analyses of the cortex, published shortly before by Brodmann (1909). He points out that in general, the myeloarchitectonic approach is superior to the cytoarchitectonic one, because the number of cortical areas that can be delineated with the aid of the former, far exceeds that delineable with the aid of the latter. Unfortunately, he does not address the specific relationship between the results of his myeloarchitectonic, and Brodmann’s cytoarchitectonic parcellation of the frontal lobe. O. Vogt (1910b) also points out that the relation between the sulci, separating the various convolutions, and the myeloarchitectonic boundaries, is variable in the frontal lobe. Some of the sulci coincide with such boundaries, but many others do not.
C. and O. Vogt (1919) published pictures of the myeloarchitecture of a number of frontal areas, some of which are reproduced in Fig. 3.11.
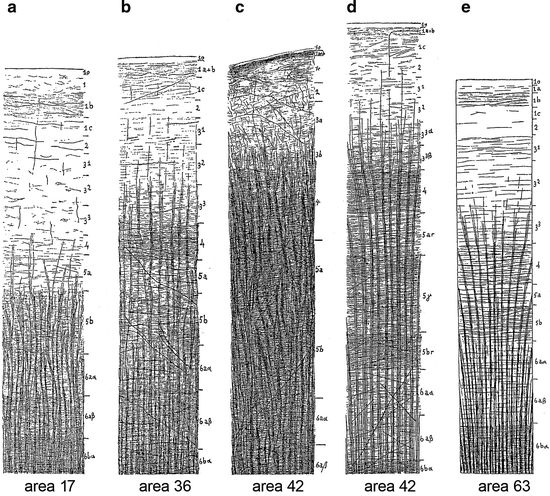
Fig. 3.11
The myeloarchitecture of some frontal areas, as depicted by C. and O. Vogt (1919). (a) Area 17, which is situated in the rostrobasal part of the cingulate gyrus (Fig. 3.9c), is of the infraradiate type, because the radii terminate principally already at the level of the transition of layers 5a and 5b. Layer 1 is subdivided, just as in the ‘Grundschema’ (Fig. 3.3), into four sublayers, one of which (1b) contains numerous ‘Einzelfasern’. There is a paucity of both radial and tangential fibres in layers 2–4, which contrasts with the much higher fibre-density seen in layers 5b and 6. Layer 5a occupies, with regard to the density of its fibres, an intermediate position between these two sets of layers. (b) Area 36 forms part of the regio unistriata euradiata grossofibrosa (Fig. 3.9a, c: III). Because of its rich endowment of fibres it is characterized as area dives. (c) Area 42 also forms part of regio III (Fig. 3.9a, c), and corresponds to a part of the cytoarchitectonic area gigantocellularis. The radii contain numerous coarse ‘Einzelfasern’. O. Vogt (1910a, p. 230) characterized this area as “absolut astriär”. C. and O. Vogt (1919) emphasized that the marking of the layers 4- 6aβ in the figure is only based on the tracing of these various cortical levels into adjacent cortical areas with a more distinct lamination. (d) Another picture of the myeloarchitecture of area 42, based on a preparation subjected to a further differentiation (‘stärkere Entfärbung’). It will be seen that distinct local differences in the density of the horizontal fibres become manifest after this procedure. (e) Area 63, forms part of the orbitofrontal sector of the regio unitostriata (Fig. 3.9b). Although the term ‘unitostriate’ refers to the situation in which both stripes of Baillarger form together a single broad band (Fig. 3.4c), in area 63 these two stripes, forming layers 4 and 5b, are nevertheless discernable as separate formations. Area 63 is designated as area pauper (Fig. 3.9b), because of its extreme poverty in ‘Grundfasern’
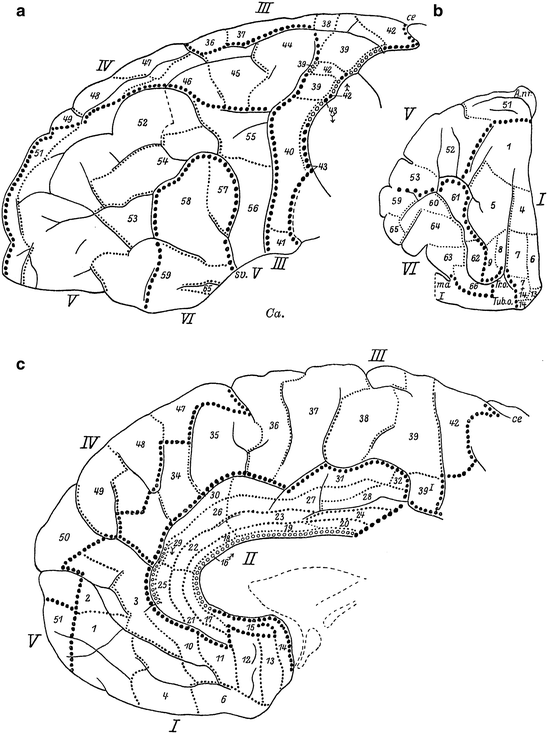
Before turning to a discussion of the publications of Strasburger (1937a, b, 1938), brief attention should be paid to Mauss’ (1908, 1911) studies on the cortex of some non-human primates. This author analyzed the myeloarchitecture of the cortex in two ‘lower’ monkeys, the rhesus macaque Macaca mulatta and the mangabey Cercocebus fulginosus (Mauss 1908), and in two anthropomorph monkeys, the gibbon Hylobatus spec. and the orang-utan Pongo pygmaeus (Mauss 1911). His principal results can be summarized as follows: (1) The results of the myeloarchtectonic analysis of the cortex of the mangabey (Fig. 3.12) closely resemble those of Brodmann’s (1905a) cytoarchitectonic analysis in the same species. (2) In the frontal lobe of the mangabey, 11 different myeloarchitectonic areas can be distinguished, nine of which (areae 4, 6, 8, 9, 10, 11, 12, and 24) can be readily equated to similarly numbered cytoarchitectonic areas. (3) The total number of delineable myeloarchitectonic cortical areas in the orang-utan (Fig. 3.13) is larger than that in the mangabey: 41 versus 31. (4) Using similarity in structure and similarity in position as criteria, 26 of the 31 myeloarchitectonic areas, present in the cortex of the mangabey, can be readily homologized with (similarly numbered) areas in the cortex of the orang-utan. (5) Although the areas numbered 8 in the maps of the mangabey and the orang-utan differ considerably in extent, they should, nevertheless, be considered as homologous. (6) The cortex of the orang-utan contains 10 myeloarchitectonic areas, which could not be identified in the mangabey. (7) All of the 11 myeloarchitectonic areas, present in the frontal lobe of the mangabey, i.e. areas 4, 6, 8, 9, 10, 11, 12, 24, 31, 32 and 33, have (similarly numbered) homologues in the frontal lobes of the orang-utan. (8) The frontal lobe of the orang-utan contains a single new myeloarchitectonic area, which is numbered 37 (Figs. 3.12 and 3.13).
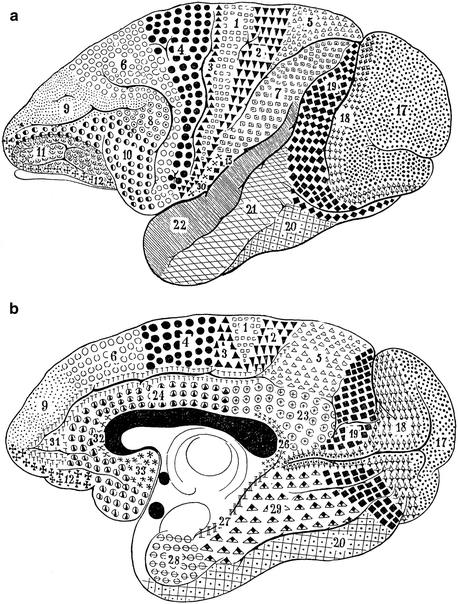
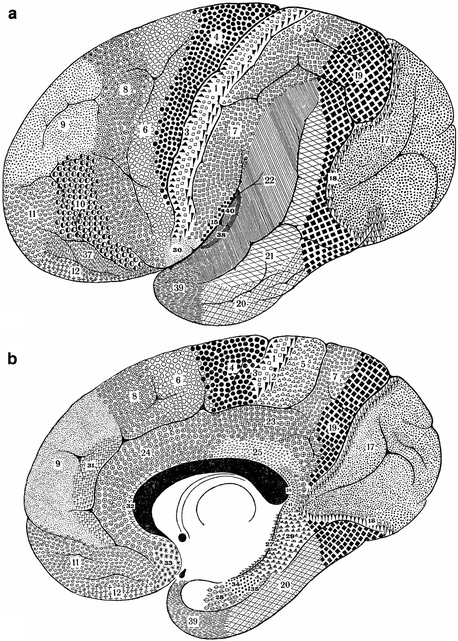
Fig. 3.12
Fig. 3.13
Mauss’ (1911) myeloarchitectonic map of the cortex of the orang-utan Pongo pygmaues; (a), lateral view; (b), medial view
Strasburger (1937a, b) thoroughly analyzed the myeloarchitecture of the frontal lobe in a human hemisphere (A 39r), and in hemispheres of two different chimpanzees (A 117l, A 118l). He compared the results of his analysis of the human frontal lobe (Fig. 3.14), with those of O. Vogt (1910a; Fig. 3.9), adopting the numbering system for the various areas of the latter; moreover, he compared the frontal lobes of the two chimpanzees studied (Fig. 3.16) with each other, as well as with that of the human (Fig. 3.14). Contrary to O. Vogt (1910a), Strasburger illustrated his descriptions of the various areas, with numerous large, very detailed drawings, some of which are reproduced (at half of their original size) in Fig. 3.15. He not only depicted individual areas, but laudably also complexes of two adjacent areas (e. g. Fig. 3.15b, c), enabling the reader to visualize the structural differences between these areas. The criteria for the establishment of homologies, used by Strasburger, were the same as those of Mauss, namely, (a) similarity in structure, and (b) similarity in position. If he was certain that a particular area in the human frontal lobe was homologous to an area delineated by O. Vogt (1910a), he adopted the number given to that area by the latter. However, if he considered the homology between a particular ‘Strasburger’ area and a particular ‘Vogt’ area as highly probable, but not entirely certain, he designated the area in question with the Roman equivalent of Vogt’s (Arabic) number of that area (cf. areas X, XI, XII and XXV in Fig. 3.14c, with areas 10, 11, 12 and 25 in Fig. 3.9c). A similar procedure was followed in the comparison of the parcellations of the frontal cortex of the two chimpanzees studied, with that of the human frontal cortex (see the numerous Roman numerals in Fig. 3.16b, d). In his characterization of the myeloarchitecture of the various areas, Strasburger paid particular attention to: (a) the disposition of the stripes of Baillarger (as regards the areas depicted in Fig. 3.15, he considered areas 51 and 61 as bistriate; areas 2, 21, 26, 30 and 50 as unistriate, and areas 64 and 65 as unitostriate); (b) the distinctness of the various sublayers in layer 1, and (c) the size and length of the radii.
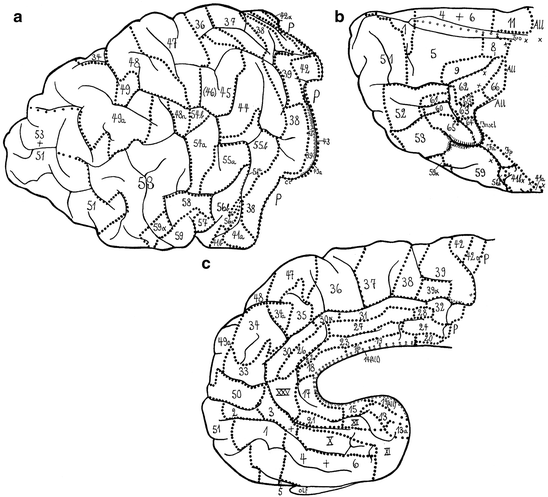
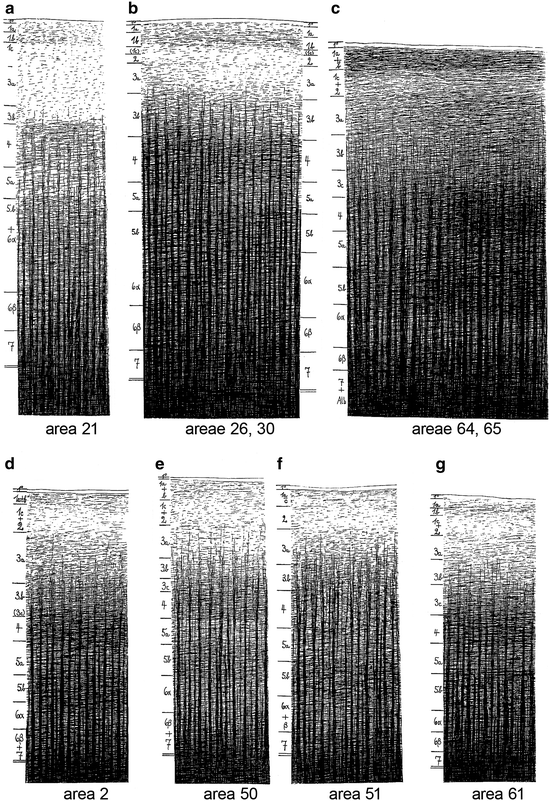
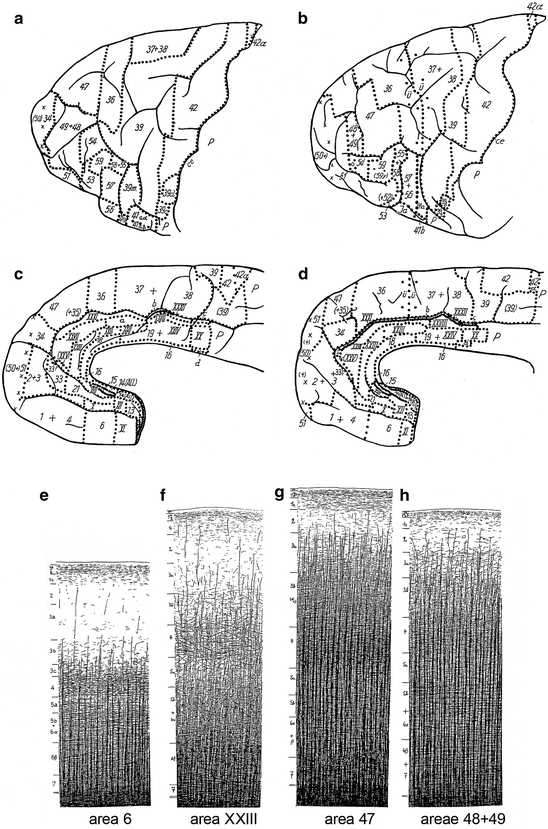
Fig. 3.14
Lateral (a), basal (b), and medial views (c) of the human frontal lobe, showing the myeloarchitectonic parcellation of Strasburger (1937a). All, allocortex; cc, central sulcus; Insel, island of Reil; olf, olfactory bulb; P, parietal lobe
Fig. 3.15
The myeloarchitecture of some areas in the human frontal lobes, as depicted by Strasburger (1937a)
Fig. 3.16
Myeloarchitectonic parcellation of the frontal cortex of the chimpanzee, according to Strasburger (1937b). (a, b) Lateral (a) and medial views (b) of the frontal lobe of specimen A 117l. (c, d) Lateral (c) and medial views (d) of the frontal lobe of specimen A 118l. (d–f) The myeloarchitecture of some fields in the frontal cortex of specimen A 117l. For abbrevaiations, see Fig. 3.11 (Reproduced from Strasburger 1937b)
The number of myeloarchitectonic areas, delineated by Strasburger in the human frontal cortex exceeds that of O. Vogt: 78 versus 66. If we disregard the slight doubts, expressed in the Roman numerals (see above), 58 of the areas distinguished by Strasburger appeared to be directly comparable to ‘Vogt-areas’. Strasburger remained unable to identify homologues of ‘Vogt-areas’ 7, 40 and 46. He split each of the five ‘Vogt-areas’ 41, 54, 55, 56 and 65, into two separate areas: 41a, 41b, 54a, 54b and so on. Finally, Strasburger distinguished 10 new areas in the frontal cortex of the human specimen studied. He did not designate these areas with new numbers. Rather, he took the number of another, adjacent area, and added the letter a, or α to it (see e. g. 49a and 59α in Fig. 3.14a).
As regards the comparison between the frontal lobes of the two chimpanzees (A 117l, A 118l), and the human studied (A 39r), Strasburger (1937b) reported that, in general, the fibre layers in the human (Fig. 3.15) are more distinct than in the chimpanzee (Fig. 3.16e–h), and that the number of areas in the human is larger: 78 versus 60 (in A 118l) or 65 (in A 117l). Some areas, including 14, 22, 26, 30, 30α, 44 and 45, present in the human frontal cortex (Fig. 3.14), could not be identified in the chimpanzee. Moreover, several sets of separate areas in the human frontal cortex, appeared to be represented by a single area (‘Sammelfeld’) in the chimpanzee (see, for example, 1+4 in Fig. 3.16b, d; 37+38 in Fig. 3.16a, c, and 48+49 in Fig. 3.16a, c, h).
Strasburger (1937b, Table 6, p. 603) compared the results of his myeloarchitectonic parcellation of the frontal cortex of the chimpanzee (50–55 areas; Fig. 3.16a–d), with those of Mauss (1911), obtained from a similar study in the orang-utan (12 areas; Fig. 3.13). He considered areas 4, 6 and 12 of Mauss directly comparable to fields 42, 39 and 6, respectively, of his parcellation. The remaining nine areas of Mauss were homologized with smaller or larger sets of areas delineated by him. To give a single example: area 11 of Mauss corresponds, according to Strasburger, with ‘his’ areas 2+3, 1+4+8, 5, 9 and 61.
In a subsequent study, Strasburger (1938) presented a detailed myeloarchitectonic analysis of frontal areas 56–66, as distinguished by O. Vogt (1910a; Fig. 3.9a, c), in both hemispheres of six human brains (A 20, A 22, A 27, A 34, A 38, A 39). With a single exception (area 63 in A 201), all of the 12 areas could be identified in all of the 12 hemispheres investigated. The 12 fields showed only very slight left-right or interspeciminal structural and positional differences. The size (volume) of some areas showed considerable differences, however.
Braitenberg’s (1956) myeloarchitectonic analysis of the human frontal cortex differs from that of O. Vogt (1910a) and Strasburger (1937a, b, 1938), in that he based his parcellation exclusively on structural differences that could be clearly observed in his Weigert-material, either with the naked eye, or with the magnifying glass. He also indicates to have included only those structural entities in his map (Fig. 3.17), that were clearly distinguishable in all of the series studied, though he does not mention on how many series his study was actually based. Another difference with the approach of O. Vogt and Strasburger is that he took the gross anatomy, i. e. the gyrification of the frontal lobe as point of departure. Thus, his frontal regions F1, F2, and F3 correspond (partly) to the superior, middle, and inferior frontal gyri, respectively, whereas his region F1 recta is centered around the straight gyrus (gyrus rectus), which forms the medial part of the basal surface of the frontal lobe. Braitenberg delineated the following myeloarchitectonic entities within the frontal lobe (Fig. 3.17):
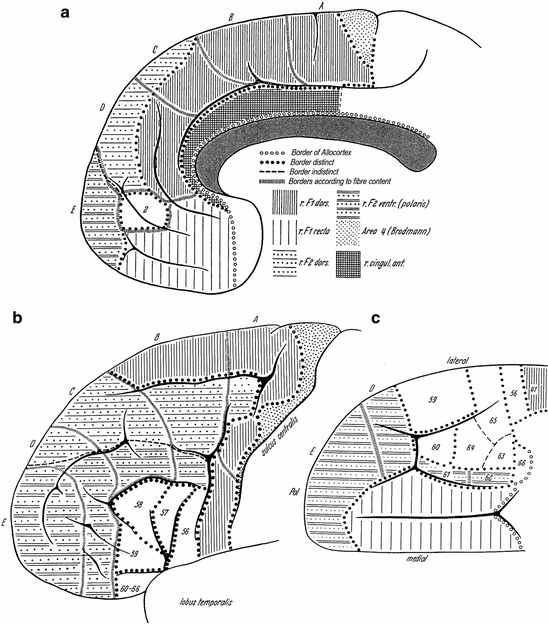
1.
An unnamed elongated, astriate region, situated directly in front of the central sulcus. This region corresponds, according to Braitenberg, with area 4 of Brodmann, and with areas 42 and 43 of O. Vogt (in what follows, all area numbers correspond, unless otherwise stated, to those of the latter).
2.
The unistriate Regio Frontalis 1 (r. F1), which includes the rostral part of the precentral gyrus, most of the superior frontal gyrus, and the medial part of the basal surface of the frontal lobe. It can be divided into a dorsal part (r. F1 dors.), and a ‘straight’ part (r. F1 recta). r. F1 dors. includes parts of O. Vogts’ regions III and IV, and encompasses areas 33–41 (cf. Fig. 3.6). r. F1 recta corresponds roughly with O. Vogts’ region I, and with the stretch of cortex occupied by areas 1–14.
3.
The bistriate Regio Frontalis 2 (r. F2), which corresponds to region V, and to areas 46–55 of O. Vogt. This region is divisible into to dorsal and ventral moieties, designated as r. F2 dors. and r. F2 vent. or – polaris. The boundary between these two divisions of r. F2 is not sharp.
4.
A small unistriate area on the medial surface of the frontal lobe, corresponding to area 2, manifesting itself, on the basis of its extraordinary wealth of fibres, clearly as a separate entity (Fig. 3.14a).
5.
The, mostly unitostriate, Regio Frontalis 3 (r. F3), which corresponds to region VI of O. Vogt, and which is clearly further differentiated into separate fields than r. F1 and r. F2. This region contains, according to O. Vogt, 11 different areas, which he numbered 56–65 (Fig. 3.9a, b). Reference to Fig. 3.17b, c shows, that Braitenberg was able to identify all of these areas, except for 61, in his material. It is of note that Strasburger (1938), as we have seen, delineated the same areas in both hemispheres of six different brains.
6.
The Regio cinguli anterior (r. cingul. ant.), which corresponds to region II of O Vogt, and which is characterized by very feebly developed stripes of Baillarger, and very short radii. Braitenberg indicates that the presence of subtle myeloarchitectonic differences, recognizable only with the aid of higher magnifications, render it possible to recognize the equivalents of areas 15–32 within the confines of this region.
Braitenberg emphasizes that the boundaries between all of the six myeloarchitectonic entities just discussed, are distinct, except for that between r. F2 dors., and r. F2 vent. However, he also points out to have observed the following changes, in passing from caudodorsal to rostroventral in r. F1 and r. F2: (i) A gradual decrease in the number of fibres; (ii) a gradual decrease in the size (calibre) of the individual fibres, and (iii) a gradual decrease in the width of the cortex. These three phenomena appeared to be correlated with (iv), a cytoarchitectonic change, viz. a gradual increase in the number of granule cells in layer IV of the cortex. These observations are noteworthy, in relation to the fact that, within the orthodox Vogt-Vogt-school (to which Braitenberg did not belong), the existence of gradual architectonic changes was categorically denied. Braitenberg made a somewhat infelicitous attempt, to indicate the gradual changes discussed, by including in his maps some borders according to fibre content (‘Grenzen nach Faserreichtum’), separating cortical compartments of ‘equal darkness’ (Fig. 3.17a–e).
In summary, Braitenberg, studying series of Weigert sections through the human frontal lobe with a simple magnifier, divided the cortex of this lobe into six large regions, r. F1 dors., r. F1 recta, r. F2 dors., r. F2 vent., r. F3, and r. cingul. ant., and two independent smaller entities, one corresponding to area 4 of Brodmann, the other to area 2 of O. Vogt. Within r. F3, eleven different areas could be delineated, almost all of which appeared to be directly comparable to myeloarchitectonic areas distinguished by O. Vogt. Passing from the central sulcus to the rostral pole of the frontal lobe, Braitenberg observed several correlated gradual structural changes in r. F1 and r. F2.
We now turn to the studies of Adolf Hopf, a collaborator of the Vogts in the Neustadt institute. Hopf’s (1956, 1968a) contributions to the myeloarchitecture of the frontal cortex are threefold. Firstly, he performed a new myeloarchitectonic parcellation of this structure; secondly, he prepared maps, showing the main myeloarchitectonic features of the frontal cortex, and thirdly, he made an attempt at the objective registration of the myeloarchitecture of the frontal lobe, using the photometric technique already discussed in a previous section.
Hopf’s (1956) renewed myeloarchitectonic parcellation of the human frontal cortex was based on serial sections of two brains, A 18, which had been previously studied by O. Vogt (1910a), and A 39, previously studied by Strasburger (1937a, b). Hopf remained, just like Strasburger, unable to distinguish areas 7 and 40, and to delimit area 45 from 46. Moreover, he regarded Strasburger’s areas 43a, 48a, 54b, 56b and 63° as inconspicuous and inconstant variants. The resultant myeloarchitectonic map, encompassing 69 areas, is shown in Fig. 3.18.
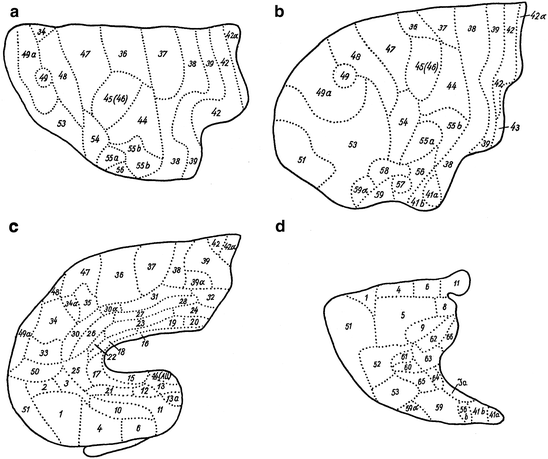
In order to visualize the distribution of the various myeloarchitectonic features over the frontal cortex, Hopf (1956) prepared separate maps, showing the overall density of fibres (Fig. 3.19), the density and size of ‘Einzelfasern’, the length of the radii, and the disposition of the stripes of Baillarger. This ‘feature-mapping’ showed, inter alia, that the precentral cortex possesses a high content of coarse fibres, and that the fibre content decreases in a step-like fashion with increasing distance from the central sulcus (Fig. 3.19).
Hopf’s (1968a) publication on the objective registration of myeloarchitectonic features in the human frontal cortex opens with a discussion of the six basic qualitative myeloarchitectonic types, occurring in this cortex (Fig. 3.20). He then presents the results of his registrations of the differential density of fibres, in stretches of cortex involving two different myeloarchitectonic areas. Two of these registrations are shown in Fig. 3.19. Hopf draws the following general conclusions from these registrations: (1) The relative fibre density of the two stripes of Baillarger in relation to each other and to the neighboring sublayers, as well as the general content in myelinated fibres, play a dominant role in these registrations. (2) The existence and reliability of some myeloarchitectonic features can be objectively demonstrated with this new technique. (3) The existence of all of the six basic, qualitatively determined types of frontal cortex (Figs. 3.20 and 3.21) could be confirmed. It is important to note that Hopf used his new myeloarchitectonic registration technique only to substantiate his qualitatively obtained maps (Fig. 3.18), and not for the creation of new, objective ‘supermaps’.
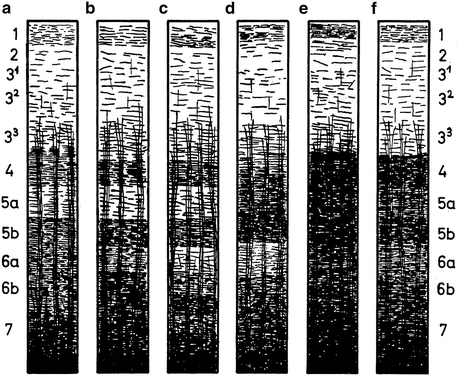
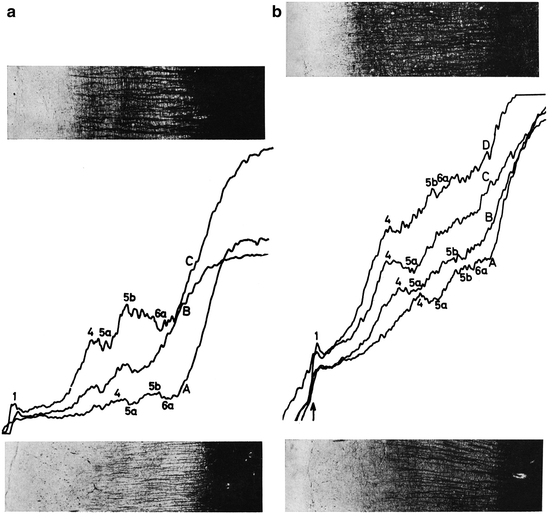
Fig. 3.20
Semidiagrammatic representation of the myeloarchitectonic types of cortex, found in the human frontal lobe. (a) Unistrate type, in which only the outer stripe of Baillarger (in layer 4) is clearly visible. (b) Propeunisstriate (= nearly unistriate) type, in which the inner stripe of B. (in layer 5b) is somewhat denser than layer 6a. (c) Bistriate type, in which both stripes of B. are well demarcated. (d) Unitostriate type, in which the two stripes of B. are united by a dense fibre plexus in layer 5a. (e) Astriate type, showing a homogeneous fibre density throughout layers 4–7. (f) Propeastriate (= nearly astriate) type, in which the stripes of B. are inconspicuous (Reproduced from Hopf 1968a)
Fig. 3.21
Photometric recordings of the fibre density in bistriate (a), and unistriate areas (b) of the human frontal cortex. The recordings in (a) are taken from area 59, shown in the lower photograph (curve A), and from area 56b, shown in the upper photograph (curve B). The recordings in (b) are taken from area 35, shown in the lower photograph (curve A), area 37 (curve B), area 38 (curve C), and area 39, shown in the upper photograph (curve D). Note that there is a clear increase in fibre content from A to D (Reproduced from Hopf 1968a)
The last study on the human frontal lobe, to be discussed here, is that of Sanides (1962, 1964), another pupil of the Vogts. This study was principally based on transversely cut serial sections through the left hemisphere of one brain, A43. The sections were stained alternately according to Nissl, for cell bodies, and according to Heidenhain-Woelcke, for myelinated fibres. Sections of other brains, including A43 and A63, were used for comparison. Sanides’ purpose was twofold: (1) To carry out a combined cytoarchitectectonic and myeloarchitectonic analysis of the frontal cortex, hence his choice of material, and (2) to work out a concept, briefly mentioned by C. and O. Vogt (1919, p. 396), according to which the architecture of the cortex shows ‘gradations’, i. e. discontinuous, stepwise changes of architectonic features.
Related posts:
 Where Matters: New Approaches to Brain Analysis
Where Matters: New Approaches to Brain Analysis
 Visualizing Myeloarchitecture In Vivo with Magnetic Resonance Imaging in Common Marmosets (Callithrix jacchus)
Visualizing Myeloarchitecture In Vivo with Magnetic Resonance Imaging in Common Marmosets (Callithrix jacchus)
 The Cytoarchitectonic Map of Constantin von Economo and Georg N. Koskinas
The Cytoarchitectonic Map of Constantin von Economo and Georg N. Koskinas
 High-Field Magnetic Resonance Mapping of the Border Between Primary Motor (Area 4) and Somatosensory (Area 3a) Cortex in Ex-Vivo and In-Vivo Human Brains
High-Field Magnetic Resonance Mapping of the Border Between Primary Motor (Area 4) and Somatosensory (Area 3a) Cortex in Ex-Vivo and In-Vivo Human Brains
 MRI Methods for In-Vivo Cortical Parcellation
MRI Methods for In-Vivo Cortical Parcellation
 Estimating the Location of Brodmann Areas from Cortical Folding Patterns Using Histology and Ex Vivo MRI
Estimating the Location of Brodmann Areas from Cortical Folding Patterns Using Histology and Ex Vivo MRI
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


