rhythm follows a circadian rhythm and that there is evidence of sleep rebound after a period of deprivation (3,4).
TABLE 3-1 HIGHLIGHTS OF IMPORTANT NEURAL CONTROL CENTERS FOR SLEEP AND WAKEFULNESS | |||||
|---|---|---|---|---|---|
|
TABLE 3-2 CHARACTERISTICS OF NREM SLEEP STAGES | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||
the 90 minutes recorded for humans (5). For example, it is about 28 minutes in cats and 10 to 12 minutes in rats (2). Humans tend to sleep during one phase in the day, whereas many mammals sleep in a “polyphasic” manner. For example, although the cat obtains a large portion of its sleep during the night, it can certainly be observed napping frequently during the daytime.
TABLE 3-3 COMPONENTS OF REM SLEEP | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
drives locus caeruleus (LC) neurons has been located (24). The SCN also influences melatonin and body temperature cycles directly, independent of dorsomedial hypothalamus contributions.
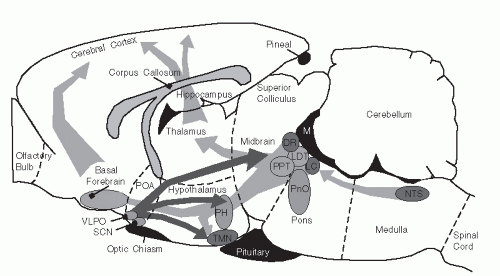 FIGURE 3-1 Schematic of location of sleep- and wake-promoting regions in the brain. Sagittal view of a rat brain redrawn from Paxinos and Watson’s The Rat Brain in Stereotaxic Coordinates (16) showing the location of major brain regions involved in wakefulness and sleep behavior. Monoaminergic nuclei are shown as dark-shaded circles. Cholinergic nuclei include the PPT and LDT nuclei. Other important nuclei are shown as light-shaded circles. Major ascending excitatory, wake-promoting projection pathways are illustrated by gray arrows, including the NTS, LC, LDT nucleus, PPT nucleus, DR, oral pontine region (PnO), the PH, including orexigenic neurons in the lateral hypothalamus, the TMN, and the SCN. The primary sleep-promoting site is located in the VLPO. The VLPO contains inhibitory neurons that project and suppress activity in wake-promoting regions. |
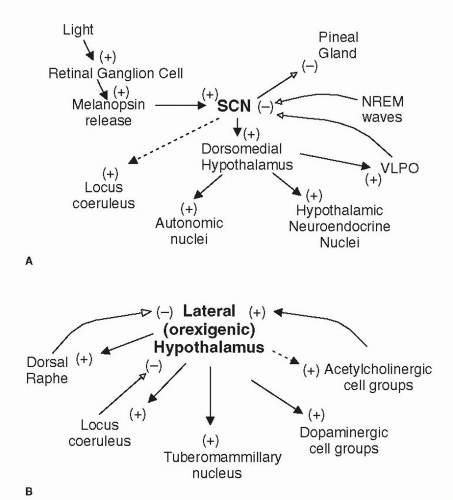 FIGURE 3-2 Interconnections of wake-promoting brain regions. (A) Influence of SCN on sleep-associated centers, autonomic, and hormonal control centers. (B) Influence of lateral hypothalamus on wake-promoting regions in the brain stem and forebrain. Solid lines indicate strong interconnections. Dashed lines indicate weak interconnections. Filled arrows and (+) represent excitatory connections. Open arrows and (-) represent inhibitory connections. |
the parabrachial nucleus, and nucleus of the solitary tract (NTS) in the dorsal medulla. Many brain stem reticular formation regions, in turn, send ascending projections back to the TMN (Fig. 3-1). The LC and ventral tegmental regions, however, send few fibers to the TMN. TMN neurons also receive afferent input from the GABAergic neurons in the VLPO, which appear to contribute strongly to the firing rate of these histaminergic neurons in relation to behavioral state.
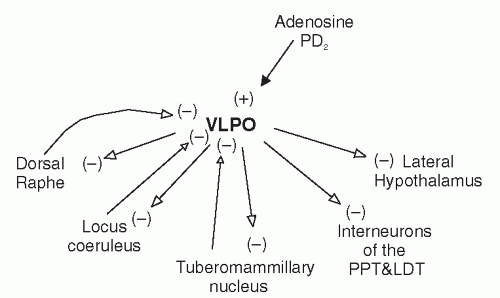 FIGURE 3-3 Interconnections of sleep-promoting brain regions. Projection sites of the inhibitory neurons in the VLPO are important in promoting sleep. VLPO are activated by wake-promoting influences such as adenosine and prostaglandin D2. Filled arrows and (+) represent excitatory connections. Open arrows and (-) represent inhibitory connections. |
Related posts:




Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


