The Spinal Cord: Introduction
The spinal cord provides a crucial information conduit, connecting the brain with most of the body. It is the target of a number of disease processes, some of which (eg, spinal cord compression) are treatable but rapidly progressive if not treated. Failure to diagnose some disorders of the spinal cord, such as spinal cord compression, can be catastrophic and may relegate the patient to a lifetime of paralysis. A knowledge of the architecture of the spinal cord and its coverings, and of the fiber tracts and cell groups that comprise it, is essential.
Development
At about the third week of prenatal development, the ectoderm of the embryonic disk forms the neural plate, which folds at the edges into the neural tube (neuraxis). A group of cells migrates to form the neural crest, which gives rise to dorsal and autonomic ganglia, the adrenal medulla, and other structures (Fig 5–1). The middle portion of the neural tube closes first; the openings at each end close later.
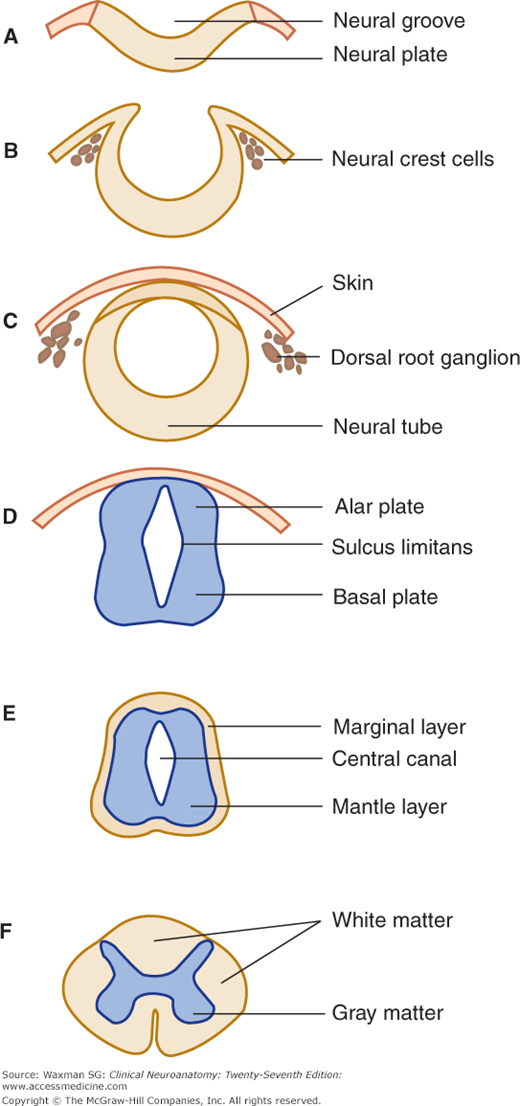
The cells in the wall of the neural tube divide and differentiate, forming an ependymal layer that encircles the central canal and is surrounded by intermediate (mantle) and marginal zones of primitive neurons and glial cells (Figs 5–1 and 5–2). The mantle zone differentiates into an alar plate, which contains mostly sensory neurons, and a basal plate, which is primarily composed of motor neurons. These two regions are demarcated by the sulcus limitans, a groove on the wall of the central canal (see Fig 5–1D). The alar plate differentiates into a dorsal gray column; the basal plate becomes a ventral gray column. The processes of the mantle zone and other cells are contained in the marginal zone, which becomes the white matter of the spinal cord (see Fig 5–2A).
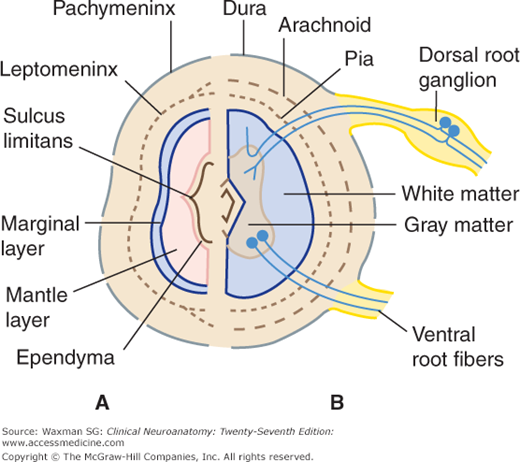
An investing layer of ectodermal cells around the primitive cord forms the two inner meninges: the arachnoid and pia mater (pia) (see Fig 5–2B). The thicker outer investment, the dura mater (dura), is formed from mesenchyma.
External Anatomy of the Spinal Cord
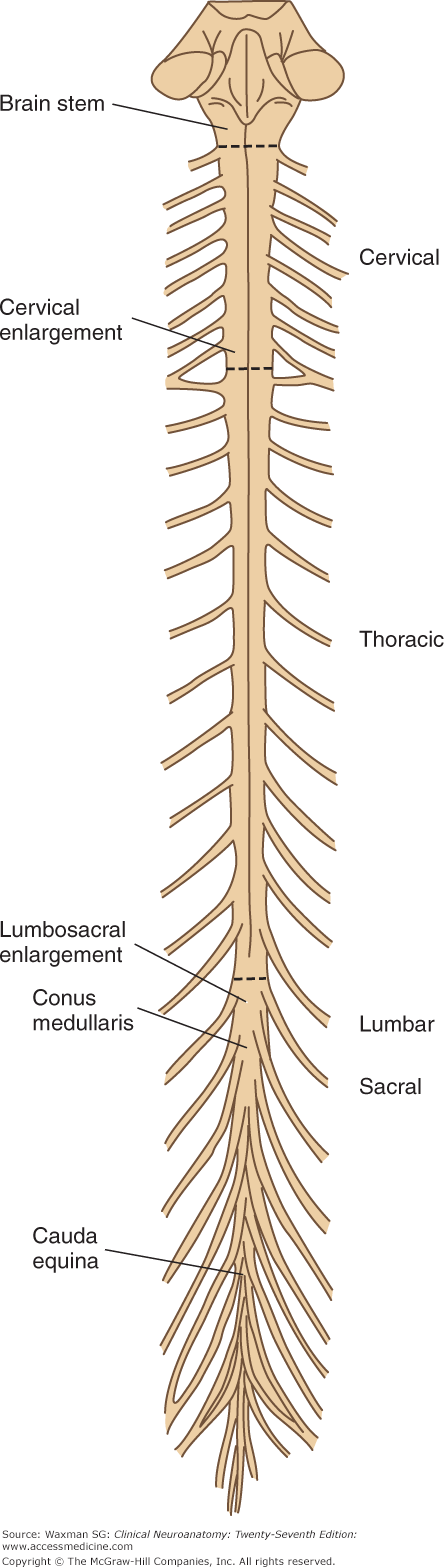
The spinal cord occupies the upper two-thirds of the adult spinal canal within the vertebral column (Fig 5–3). The cord is normally 42 to 45 cm long in adults and is continuous with the medulla at its upper end. The conus medullaris is the conical distal (inferior) end of the spinal cord. In adults, the conus ends at the L1 or L2 level of the vertebral column. The filum terminale extends from the tip of the conus and attaches to the distal dural sac. The filum terminale consists of pia and glial fibers and often contains a vein.
The central canal is lined with ependymal cells and filled with cerebrospinal fluid. It opens upward into the inferior portion of the fourth ventricle.
The spinal cord widens laterally in the cervical enlargement and the lumbosacral enlargement (see Fig 5–3). The latter tapers off to form the conus medullaris. The enlargements of the cord contain increased numbers of lower motor neurons (LMNs) and provide the origins of the nerves of the upper and lower extremities. The nerves of the brachial plexus originate at the cervical enlargement; the nerves of the lumbosacral plexus arise from the lumbar enlargement.
The spinal cord can be thought of as consisting of approximately 30 segments (see Fig 5–3 and Appendix C)—8 cervical (C) segments, 12 thoracic (T) segments (termed dorsal in some texts), 5 lumbar (L) segments, 5 sacral (S) segments, and a few small coccygeal (Co) segments—that correspond to attachments of groups of nerve roots (Figs 5–3 and 5–4). There are no sharp boundaries between segments within the cord itself.
Figure 5–4
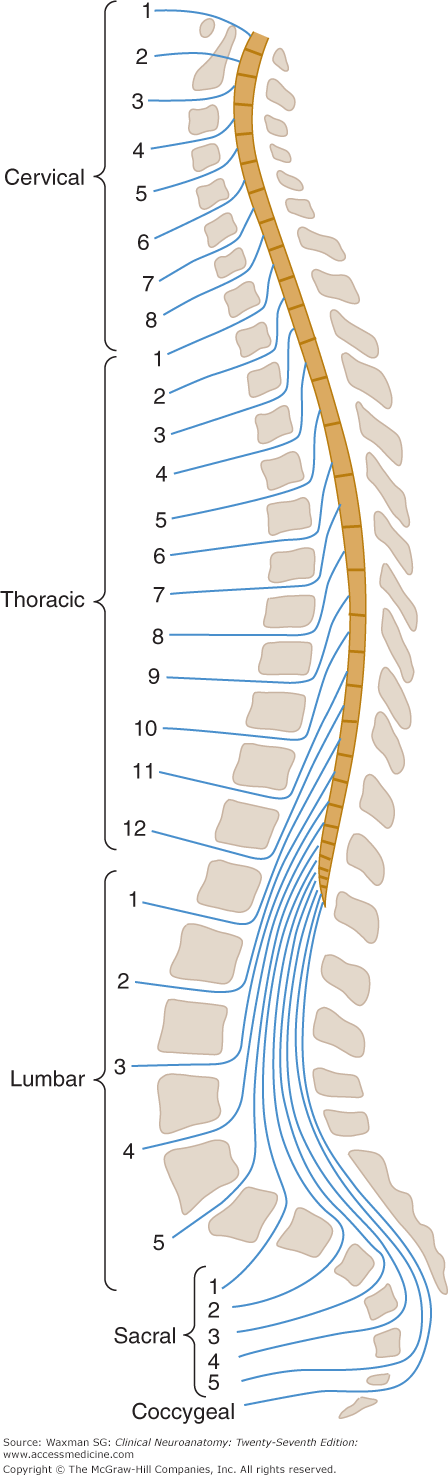
Schematic illustration of the relationships between the vertebral column, the spinal cord, and the spinal nerves. Note the mismatch between the location of spinal cord segments and of vertebral level where roots exit from the vertebral column. Note also the termination of the spinal cord at the level of the L1 or L2 vertebral body.
Because the spinal cord is shorter than the vertebral column, each spinal cord segment at lower levels is located above the similarly numbered vertebral body. The relation between spinal cord segments and vertebral bodies is shown in Table 5–1 and Figure 5–4.
A cross section of the spinal cord shows a deep anterior median fissure and a shallow posterior (or dorsal) median sulcus, which divide the cord into symmetric right and left halves joined in the central midportion (Fig 5–5). The anterior median fissure contains a fold of pia and blood vessels; its floor is the anterior (or ventral) white commissure. The dorsal nerve roots are attached to the spinal cord along a shallow vertical groove, the posterolateral sulcus, which lies at a short distance anterior to the posterior median sulcus. The ventral nerve roots exit in the anterolateral sulcus.
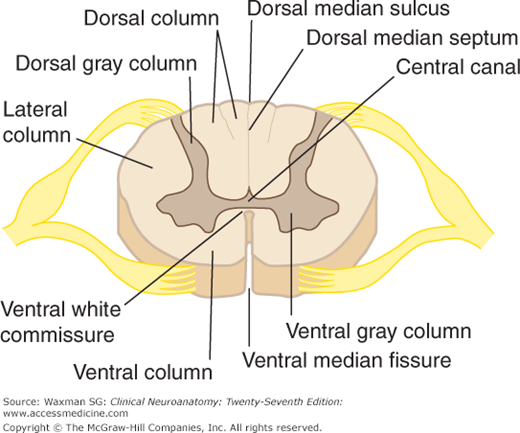
A note on terminology: In descriptions of the spinal cord, the terms ventral and anterior are used interchangeably. Similarly, dorsal and posterior have the same meaning when referring to the spinal cord and its tracts; the dorsal columns, for example, are sometimes referred to as the posterior columns.
Clinical Correlations
Failure of the neural tube to close at the cranial end results in anencephaly, a type of maldevelopment of the brain and skull that is incompatible with life. Failure of closure at the caudal end results in spina bifida, which is associated with maldevelopment of the vertebrae (see Fig 6–9).
Spinal Roots and Nerves
Each segment of the spinal cord gives rise to four roots: a ventral and a dorsal root on the left and a similar pair on the right (see Fig 5–5). The first cervical segment usually lacks dorsal roots.
Each of the 31 pairs of spinal nerves has a ventral root and a dorsal root; each root is made up of 1 to 8 rootlets (Fig 5–6). Each root consists of bundles of nerve fibers. In the dorsal root of a typical spinal nerve, close to the junction with the ventral root, lies a dorsal root (spinal) ganglion, a swelling that contains nerve cell bodies that give rise to sensory axons. The portion of a spinal nerve outside the vertebral column is sometimes referred to as a peripheral nerve. The spinal nerves are divided into groups that correspond to the spinal cord segments (see Fig 5–4).
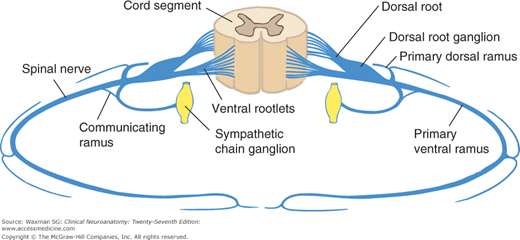
The vertebral column surrounds and protects the spinal cord and normally consists of 7 cervical, 12 thoracic, and 5 lumbar vertebrae as well as the sacrum, which is usually formed by fusion of 5 vertebrae, and the coccyx. The nerve roots exit from the vertebral column through intervertebral foramina. In the cervical spine, the numbered roots exit the vertebral column above the corresponding vertebral body. The C8 root exits between vertebral bodies C7 and T1. In the lower parts of the spine, the numbered roots exit below the correspondingly numbered vertebral body.
The spinal cord itself is shorter than the vertebral column, and it usually ends at L1–2. The anatomy of the vertebral column is discussed further in Chapter 6.
Until the third month of fetal life, the spinal cord is as long as the vertebral canal. After that point, the vertebral column elongates faster than the spinal cord, so that at birth the cord extends to about the level of the third lumbar vertebra. In adults, the tip of the cord normally lies at the level of the first or second lumbar vertebra. Because of the different growth rates of the cord and spine, the cord segments are displaced upward from their corresponding vertebrae, with the greatest discrepancy in the lowest segments (see Fig 5–4). In the lumbosacral region, the nerve roots descend almost vertically below the cord to form the cauda equina (horse’s tail) (see Figs 5–3 and 5–4).
The ventral (or anterior) roots constitute motor outflow tracts from the spinal cord. The ventral roots carry the large-diameter alpha motor neuron axons to the extrafusal striated muscle fibers; the smaller gamma motor neuron axons, which supply the intrafusal muscle of the muscle spindles (Fig 5–7); preganglionic autonomic fibers at the thoracic, upper lumbar, and midsacral levels (see Chapter 20); and a few afferent, small-diameter axons that arise from cells in the dorsal root ganglia and convey sensory information from the thoracic and abdominal viscera.
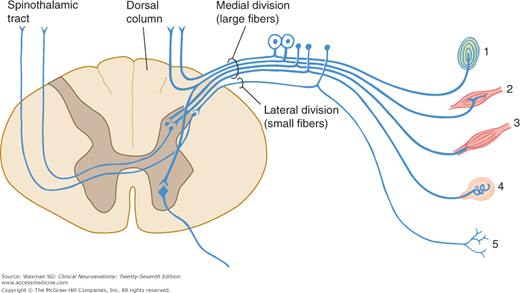
The dorsal (posterior) roots are largely sensory. Each dorsal nerve root (except usually C1) contains afferent fibers from the nerve cells in its ganglion. The dorsal roots contain fibers from cutaneous and deep structures (see Table 3–2). The largest fibers (Ia) come from muscle spindles and participate in spinal cord reflexes; the medium-sized fibers (A-beta) convey impulses from mechanoreceptors in skin and joints. Most of the axons in the dorsal nerve roots are small (C, nonmyelinated; A-delta, myelinated) and carry information of noxious (eg, pain) and thermal stimuli.
This consists of a medial branch, which is in most instances largely sensory, and a lateral branch, which is mainly motor.
Larger than the posterior primary division, the anterior primary divisions form the cervical, brachial, and lumbosacral plexuses. In the thoracic region they remain segmental, as intercostal nerves.
The rami join the spinal nerves to the sympathetic trunk. Only the thoracic and upper lumbar nerves contain a white ramus communicans, but the gray ramus is present in all spinal nerves (see Fig 5–6).
These nerves, also called sinuvertebral nerves, are quite small; they carry sensory and vasomotor innervation to the meninges.
Nerve fibers can be classified not only on the basis of their diameter and conduction velocity (see Tables 3–2 and 3–3) but also on a physioanatomic basis.
These motor fibers innervate the skeletal muscles. They originate in large cells in the anterior gray column of the spinal cord and form the ventral root of the spinal nerve.
These fibers convey sensory information from the skin, joints, and muscles to the central nervous system. Their cell bodies are unipolar cells in the spinal ganglia that are interposed in the course of dorsal roots (dorsal root ganglia). The peripheral branches of these ganglionic cells are distributed to somatic structures; the central branches convey sensory impulses through the dorsal roots to the dorsal gray column and the ascending tracts of the spinal cord.
The autonomic fibers are the motor fibers to the viscera. Sympathetic fibers from the thoracic segments and L1 and L2 are distributed throughout the body to the viscera, glands, and smooth muscle. Parasympathetic fibers, which are present in the middle three sacral nerves, go to the pelvic and lower abdominal viscera. (Other parasympathetic fibers are carried by cranial nerves III, VII, IX, and X.)
These fibers convey sensory information from the viscera. Their cell bodies are in the dorsal root ganglia.
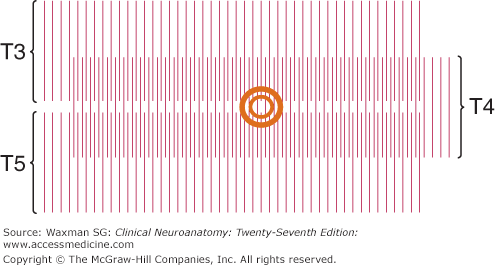
The sensory component of each spinal nerve is distributed to a dermatome, a well-defined segmental portion of the skin (Fig 5–8). An understanding of the dermatomes is essential for interpretion of the sensory examination. It is important for all clinicians to remember the following key points:
- Because in many patients there is no C1 dorsal root, there is no C1 dermatome (when a C1 dermatome does exist as an anatomic variant, it covers a small area in the central part of the neck, close to the occiput).
- The dermatomes for C5, C6, C7, C8, and T1 are confined to the arm, and the C4 and T2 dermatomes are contiguous over the anterior trunk.
- The thumb, middle finger, and fifth digit are within the C6, C7, and C8 dermatomes, respectively.
- The nipple is at the level of T4.
- The umbilicus is at the level of T10.
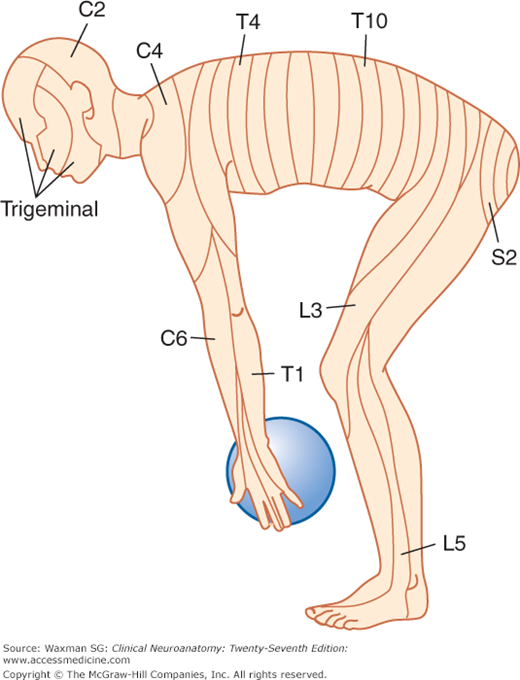
The territories of dermatomes tend to overlap, making it difficult to determine the absence of a single segmental innervation on the basis of sensory testing (Fig 5–9).
The term myotome refers to the skeletal musculature innervated by motor axons in a given spinal root. The organization of myotomes is the same from person to person, and the testing of motor functions (see Appendix B) can be very useful in determining the extent of a lesion in the nerve, spinal cord segment, or tract, especially when combined with a careful sensory examination. Most muscles, as indicated in Appendix B, are innervated by motor axons that arise from several adjacent spinal roots. Nevertheless, lesions of a single spinal root, in many cases, can cause weakness and atrophy of a muscle. Table 5–2 lists segment-pointer muscles, whose weakness or atrophy may suggest a lesion involving a single nerve root or a pair of adjacent nerve roots.
Muscle | Spinal Root | Function of Muscle |
|---|---|---|
Diaphragm | C3, C4 | Respiration |
Deltoid | C5 | Arm Abduction |
Biceps | C5 | Forearm Flexion |
Brachioradialis | C6 | Forearm Flexion |
Triceps | C7 | Extension of Forearm |
Quadriceps femoris | L3, L4 | Knee Extension |
Tibialis Anterior | L4 | Dorsiflexion of Foot |
Extensor hallucis longus | L5 | Dorsiflexion of Great Toe |
Gastrocnemius | S1 | Plantar Flexion |
Internal Divisions of the Spinal Cord
A cross section of the spinal cord shows an H-shaped internal mass of gray matter surrounded by white matter (see Fig 5–5). The gray matter is made up of two symmetric portions joined across the midline by a transverse connection (commissure) of gray matter that contains the minute central canal or its remnants. This gray matter extends up and down the entire length of the spinal cord, and is considered to consist of columns. The ventral (or anterior) gray column (also called the ventral, or anterior, horn) is in front of the central canal. It contains the cells of origin of the fibers of the ventral roots, including alpha and gamma motor neurons (“lower” motor neurons).
The intermediolateral gray column (or horn) is the portion of gray matter between the dorsal and ventral gray columns; it is a prominent lateral triangular projection in the thoracic and upper lumbar regions but not in the midsacral region. It contains preganglionic cells for the autonomic nervous system. Within spinal segments T1 to L2, preganglionic sympathetic neurons within the intermediolateral gray column give rise to sympathetic axons that leave the spinal cord within the ventral roots and then travel to the sympathetic ganglia via the white rami communicantes. Within spinal segments S2, S3, and S4, there are sacral parasympathetic neurons within the intermediolateral gray column. These neurons give rise to preganglionic parasympathetic axons that leave the spinal cord within the sacral ventral roots. After projecting to the pelvic viscera within the pelvic nerves, these parasympathetic axons synapse on postganglionic parasympathetic neurons that project to the pelvic viscera.
The dorsal gray column (also called the posterior, or dorsal, horn) reaches almost to the posterolateral (dorsolateral) sulcus. A compact bundle of small fibers, the dorsolateral fasciculus (Lissauer’s tract), part of the pain pathway, lies on the periphery of the spinal cord.
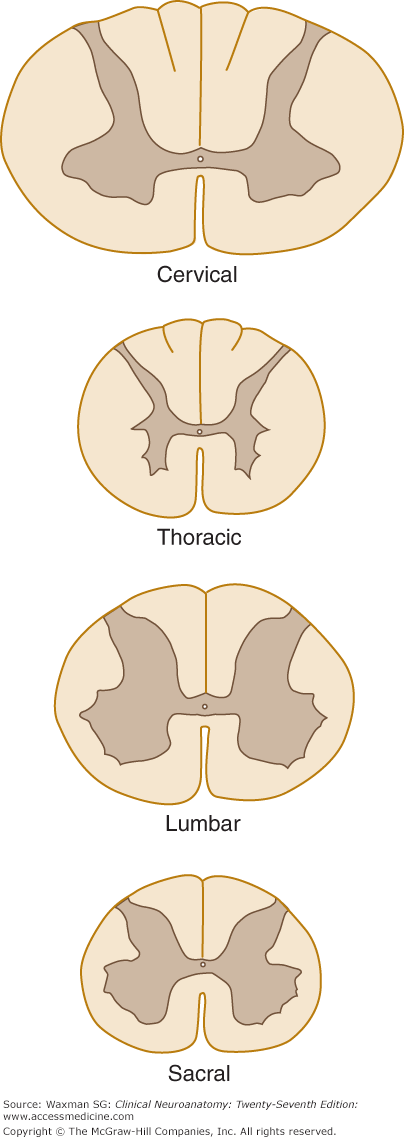
The form and quantity of the gray matter vary at different levels of the spinal cord (Fig 5–10). The proportion of gray to white matter is greatest in the lumbar and cervical enlargements. In the cervical region, the dorsal gray column is comparatively narrow and the ventral column is broad and expansive, especially in the four lower cervical segments. In the thoracic region, both the dorsal and ventral columns are narrow, and there is a lateral column. In the lumbar region, the dorsal and ventral columns are broad and expanded. In the conus medullaris, the gray matter looks like two oval masses, one in each half of the cord, connected by a wide gray commissure.
A cross section of the gray matter of the spinal cord shows a number of laminas (layers of nerve cells), termed Rexed’s laminae after the neuroanatomist who described them (Fig 5–11). As a general principle, superficial laminae tend to be involved in pain signaling, while deeper laminae are involved in non-painful as well as painful sensation.
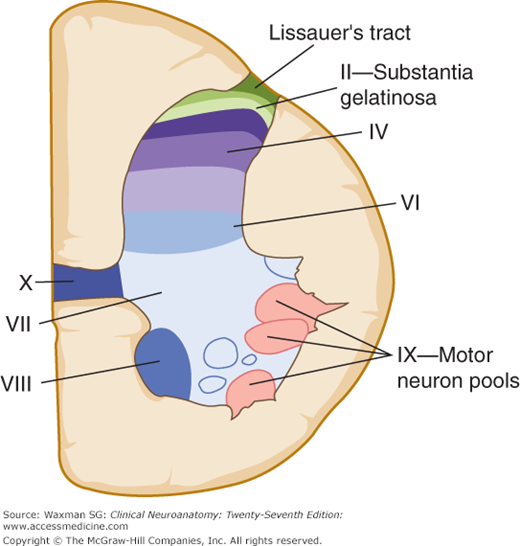
This thin marginal layer contains neurons that respond to noxious stimuli and send axons to the contralateral spinothalamic tract.
Also known as substantia gelatinosa, this lamina is made up of small neurons, some of which respond to noxious stimuli. Substance P, a neuropeptide involved in pathways mediating sensibility to pain, is found in high concentrations in laminas I and II.
These are referred to together as the nucleus proprius. Their main input is from fibers that convey position and light touch sense.
This layer contains cells that respond to both noxious and visceral afferent stimuli.
This deepest layer of the dorsal horn contains neurons that respond to mechanical signals from joints and skin.
This is a large zone that contains the cells of the dorsal nucleus (Clarke’s column) medially as well as a large portion of the ventral gray column. Clarke’s column contains cells that give rise to the posterior spinocerebellar tract. Lamina VII also contains the intermediolateral nucleus (or intermediolateral cell column) in thoracic and upper lumbar regions. Preganglionic sympathetic fibers project from cells in this nucleus, via the ventral roots and white rami communicantes, to sympathetic ganglia.
These layers represent motor neuron groups in the medial and lateral portions of the ventral gray column. The medial portion (also termed the medial motor neuron column) contains the LMNs that innervate axial musculature (ie, muscles of the trunk and proximal parts of the limbs). The lateral motor neuron column contains LMNs for the distal muscles of the arm and leg. In general, flexor muscles are innervated by motor neurons located close to the central canal, whereas extensor muscles are innervated by motor neurons located more peripherally (Fig 5–12).
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


