Ventricular System
Within the brain is a communicating system of cavities that are lined with ependyma and filled with cerebrospinal fluid (CSF): There are two lateral ventricles, the third ventricle (between the halves of the diencephalon), the cerebral aqueduct, and the fourth ventricle within the brain stem (Fig 11–1).
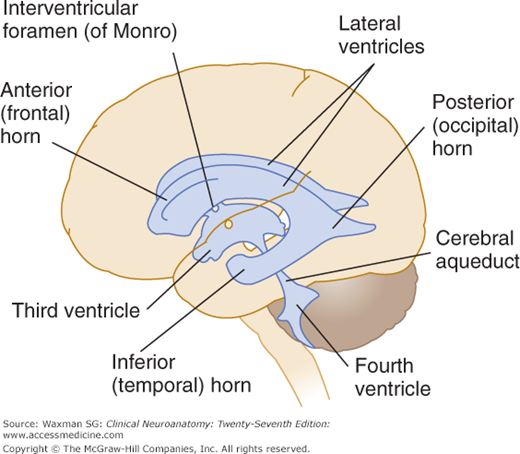
The lateral ventricles are the largest. They each include two central portions (body and atrium) and three extensions (horns).
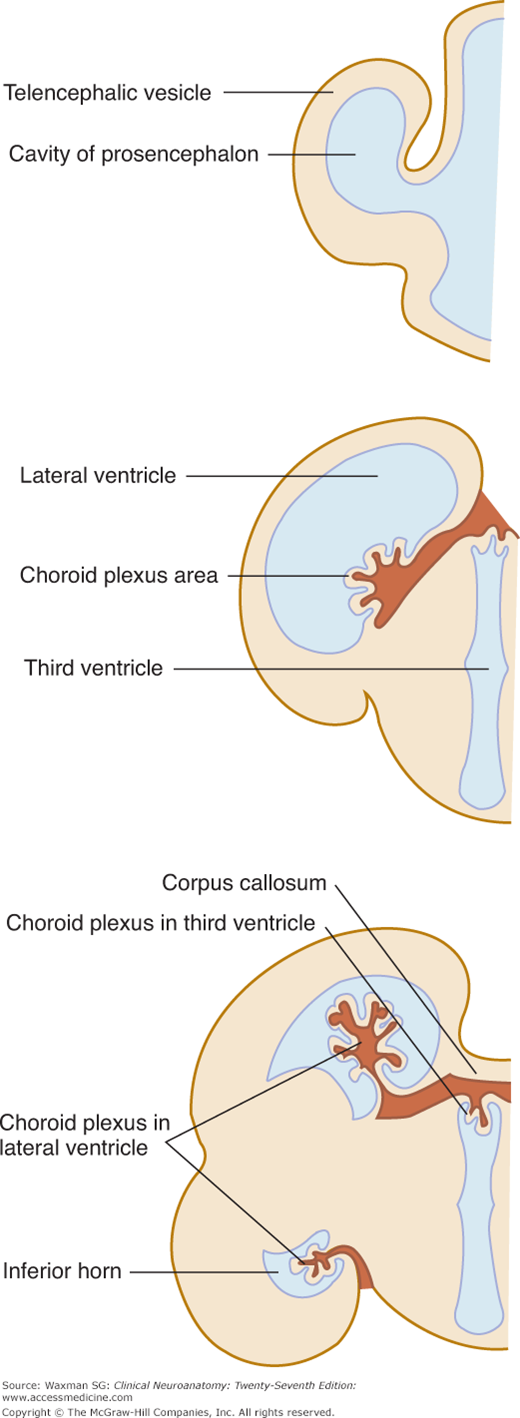
The choroid plexus is the site where cerebrospinal fluid (CSF) is produced. It is a fringe-like vascular process of pia mater containing capillaries of the choroid arteries. It projects into the ventricular cavity and is covered by an epithelial layer of ependymal origin (Figs 11–2 and 11–3). The attachment of the plexus to the adjacent brain structures is known as the tela choroidea. The choroid plexus extends from the interventricular foramen, where it joins with the plexuses of the third ventricle and opposite lateral ventricle, to the end of the inferior horn. (There is no choroid plexus in the anterior and posterior horns.)
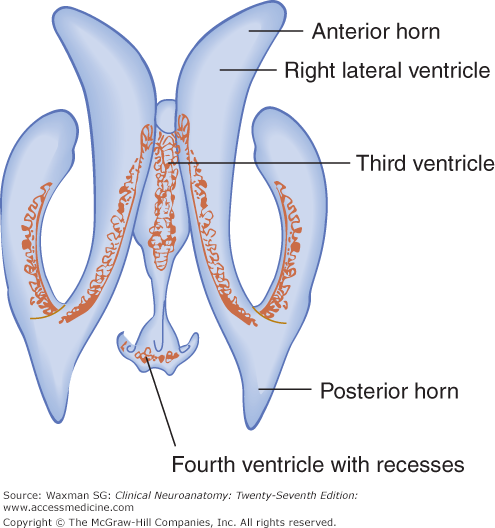
The anterior (frontal) horn is in front of the interventricular foramen. Its roof and anterior border are formed by the corpus callosum; its vertical medial wall, by the septum pellucidum; and the floor and lateral wall, by the bulging head of the caudate nucleus.
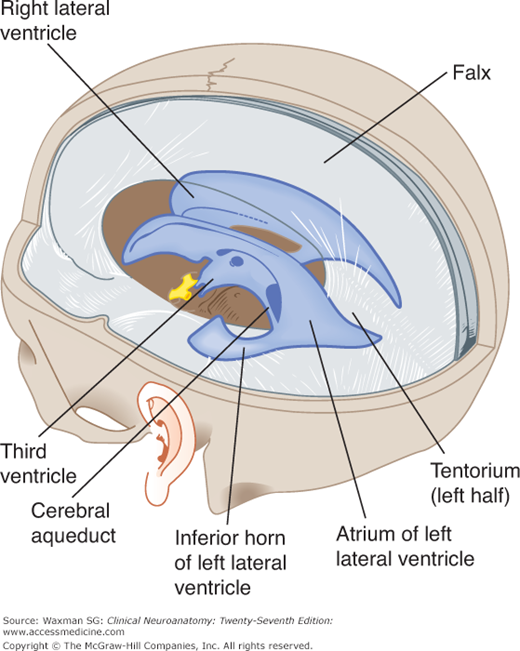
The central part, or body, of the lateral ventricle is the long, narrow portion that extends from the interventricular foramen to a point opposite the splenium of the corpus callosum. Its roof is formed by the corpus callosum and its medial wall by the posterior portion of the septum pellucidum. The floor contains (from medial to lateral side) the fornix, the choroid plexus, the lateral part of the dorsal surface of the thalamus, the stria terminalis, the vena terminalis, and the caudate nucleus. The atrium, or trigone, is a wide area of the body that connects with the posterior and inferior horns (Fig 11–4).
The posterior (occipital) horn extends into the occipital lobe. Its roof is formed by fibers of the corpus callosum. On its medial wall is the calcar avis, an elevation of the ventricular wall produced by the calcarine fissure.
The inferior (temporal) horn traverses the temporal lobe, whose white substance forms its roof. Along the medial border are the stria terminalis and the tail of the caudate nucleus. The amygdaloid nuclear complex bulges into the upper terminal part of the inferior horn, whose floor and medial wall are formed by the fimbria, hippocampus, and collateral eminence.
The two interventricular foramens, or foramens of Monro, are apertures between the column of the fornix and the anterior end of the thalamus. The two lateral ventricles communicate with the third ventricle through these foramens (see Fig 11–1).
The third ventricle is a narrow vertical cleft between the two halves of the diencephalon (see Figs 11–1, 11–2, 11–3, and 11–4). The roof of the third ventricle is formed by a thin tela choroidea (a layer of ependyma) and pia mater from which a small choroid plexus extends into the lumen of the ventricle (see Fig 9–1). The lateral walls are formed mainly by the medial surfaces of the two thalami. The lower lateral wall and the floor of the ventricle are formed by the hypothalamus; the anterior commissure and the lamina terminalis form the rostral limit.
The optic recess is an extension of the third ventricle between the lamina terminalis and the optic chiasm. The hypophysis is attached to the apex of its downward extension, the funnel-shaped infundibular recess. A small pineal recess projects into the stalk of the pineal body. A large extension of the third ventricle above the epithalamus is known as the suprapineal recess.
The cerebral aqueduct is a narrow, curved channel running from the posterior third ventricle into the fourth. It contains no choroid plexus (see Figs 11–1 and 11–4).
The fourth ventricle is a pyramid-shaped cavity bounded ventrally by the pons and medulla oblongata (see Figs 7–14, 11–1, and 11–3); its floor is also known as the rhomboid fossa. The lateral recess extends as a narrow, curved extension of the ventricle on the dorsal surface of the inferior cerebellar peduncle. The fourth ventricle extends under the obex into the central canal of the medulla.
The incomplete roof of the fourth ventricle is formed by the anterior and posterior medullary vela. The anterior medullary velum extends between the dorsomedial borders of the superior cerebellar peduncles, and its dorsal surface is covered by the adherent lingula of the cerebellum. The posterior medullary velum extends caudally from the cerebellum. The point at which the fourth ventricle passes up into the cerebellum is called the apex, or fastigium.
The position of the cerebellum, just above the roof of the fourth ventricle, has important clinical implications. Mass lesions of the cerebellum (eg, tumors) or swelling of the cerebellum after a cerebellar infarction can compress the fourth ventricle, producing acute obstructive hydrocephalus.
The lateral aperture (foramen of Luschka) is the opening of the lateral recess into the subarachnoid space near the flocculus of the cerebellum. A tuft of choroid plexus is commonly present in the aperture and partly obstructs the flow of CSF from the fourth ventricle to the subarachnoid space. The medial aperture (foramen of Magendie) is an opening in the caudal portion of the roof of the ventricle. Most of the outflow of CSF from the fourth ventricle passes through this aperture, which varies in size.
The tela choroidea of the fourth ventricle is a layer of pia and ependyma that contains small vessels and lies in the posterior medullary velum. It forms the choroid plexus of the fourth ventricle.
Meninges and Submeningeal Spaces
Three membranes, or meninges, envelop the brain: the dura, the arachnoid, and the pia. The dura, the outer membrane, is separated from the thin arachnoid by a potential compartment, the subdural space, which normally contains only a few drops of CSF. An extensive subarachnoid space containing CSF and the major arteries separates the arachnoid from the pia, which completely invests the brain. The arachnoid and the pia, known collectively as leptomeninges, are connected by thin strands of tissue, the arachnoid trabeculae. The pia, together with a narrow extension of the subarachnoid space, accompanies the vessels deep into the brain tissue; this space is called the perivascular space, or Virchow–Robin’s space.
The dura, which was formerly called the pachymeninx, is a tough, fibrous structure with an inner (meningeal) and an outer (periosteal) layer (Figs 11–4 and 11–5). (Most of the dura’s venous sinuses lie between the dural layers.) The dural layers over the brain are generally fused, except where they separate to provide space for the venous sinuses and where the inner layer forms septa between brain portions. The outer layer is firmly attached to the inner surface of the cranial bones and sends vascular and fibrous extensions into the bone itself; the inner layer is continuous with the spinal dura.
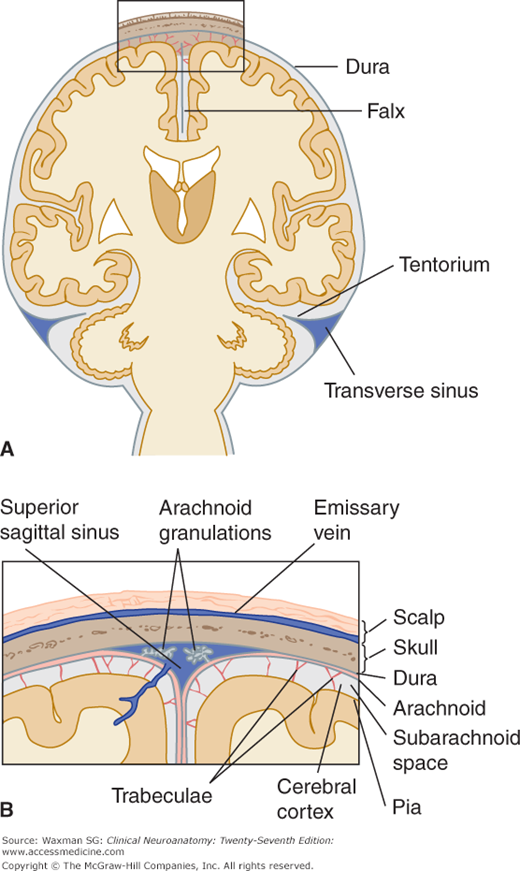
One of the dural septa, the falx cerebri, extends like a curtain, down into the longitudinal fissure between the cerebral hemispheres (Figs 11–5 and 11–6). It attaches to the inner surface of the skull in midplane, from the crista galli to the internal occipital protuberance, where it becomes continuous with the tentorium cerebelli.
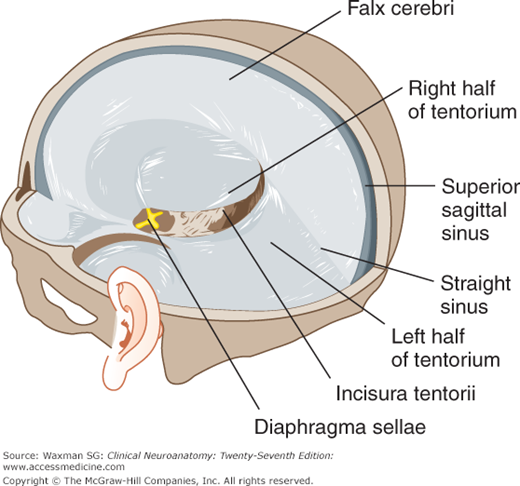
The tentorium cerebelli separates the occipital lobes from the cerebellum. It is a roughly transverse, shelflike membrane that attaches at the rear and side to the skull at the transverse sinuses; at the front, it attaches to the petrous portion of the temporal bone and to the clinoid processes of the sphenoid bone. Toward the midline, it fuses with the falx cerebri. The free, curved anterior border leaves a large opening, the incisura tentorii (tentorial notch), for passage of the upper brain stem, aqueduct, and vessels.
The falx cerebelli projects between the cerebellar hemispheres from the inner surface of the occipital bone to form a small triangular dural septum.
The diaphragma sellae forms an incomplete lid over the hypophysis in the sella turcica by connecting the clinoid attachments of the two sides of the tentorium cerebelli. The pituitary stalk passes through the opening in the diaphragma.
The arachnoid, a delicate avascular membrane, covers the subarachnoid space, which is filled with CSF. The inner surface of the arachnoid is connected to the pia by fine (but inconstantly present) arachnoid trabeculae (see Fig 11–5). The cranial arachnoid closely covers the inner surface of the dura mater but is separated from it by the subdural space, which contains a thin film of fluid. The arachnoid does not dip into the sulci or fissures except to follow the falx and the tentorium.
Arachnoid granulations consist of many microscopic villi (see Fig 11–5B). They have the appearance of berry-like clumps protruding into the superior sagittal sinus or its associated venous lacunae and into other sinuses and large veins. The granulations are sites of absorption of CSF.
The subarachnoid space between the arachnoid and the pia is relatively narrow over the surface of the cerebral hemisphere, but it becomes much wider in areas at the base of the brain. These widened spaces, the subarachnoid cisterns, are often named after neighboring brain structures (Fig 11–7). They communicate freely with adjacent cisterns and the general subarachnoid space.

The cisterna magna results from the bridging of the arachnoid over the space between the medulla and the cerebellar hemispheres; it is continuous with the spinal subarachnoid space. The pontine cistern on the ventral aspect of the pons contains the basilar artery and some veins. Below the cerebrum lies a wide space between the two temporal lobes. This space is divided into the chiasmatic cisterna above the optic chiasm, the suprasellar cistern above the diaphragma sellae, and the interpeduncular cistern between the cerebral peduncles. The space between frontal, parietal, and temporal lobes is called the cistern of the lateral fissure (cistern of Sylvius).
The pia is a thin connective tissue membrane that covers the brain surface and extends into sulci and fissures and around blood vessels throughout the brain (see Fig 11–5). It also extends into the transverse cerebral fissure under the corpus callosum. There it forms the tela choroidea of the third and lateral ventricles and combines with the ependyma and choroid vessels to form the choroid plexus of these ventricles. The pia and ependyma pass over the roof of the fourth ventricle and form its tela choroidea for the choroid plexus there.
Clinical Correlations
Several types of herniation of the brain can occur (Fig 11–8). The tentorium separates the supratentorial and the infratentorial compartments, and the two spaces communicate by way of the incisura that contains the midbrain. Both the falx and the tentorium form incomplete separations, and a mass or expanding lesion may displace a portion of the brain around these septa, resulting in either a subfalcial or a transtentorial herniation. In subfalcial herniation, the cingulate gyrus is displaced into or under the falx. In transtentorial herniation, the uncus (of the medial temporal lobe) is displaced through the tentorium, and compresses the brain stem and the adjacent oculomotor nerve (causing an ipsilateral dilated pupil and third nerve paresis). Herniation of the cerebellar tonsils into the foramen magnum by a lesion is often called coning. Transtentorial and cerebellar tonsillar herniation are life threatening because they can distort or compress the brain stem and damage its vital regulatory centers for respiration, consciousness, blood pressure, and other functions (see Chapters 18 and 20).
Figure 11–8

Anatomic basis of herniation syndromes. An expanding supratentorial mass lesion may cause brain tissue to be displaced into an adjacent intracranial compartment, resulting in (1) cingulate herniation under the falx, (2) downward transtentorial (central) herniation, (3) uncal herniation over the edge of the tentorium, or (4) cerebellar tonsillar herniation into the foramen magnum. Coma and ultimately death result when (2), (3), or (4) produces brainstem compression. (Reproduced with permission from Aminoff ML, Greenberg DA, Simon RP: Clinical Neurology, 6th edition, McGraw-Hill, 2005.)
CSF
The CSF acts like a protective water jacket around the brain. It controls brain excitability by regulating the ionic composition, carries away metabolites (because the brain has no lymphatic vessels), and provides protection from pressure changes (venous volume versus CSF volume).









