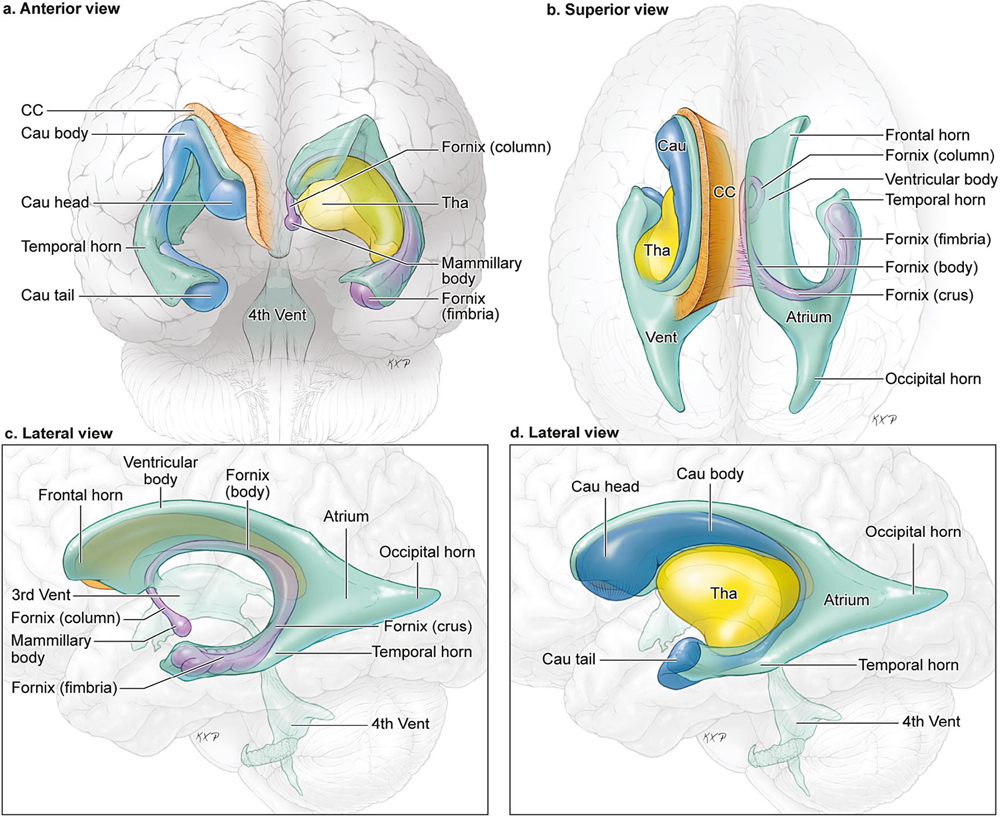
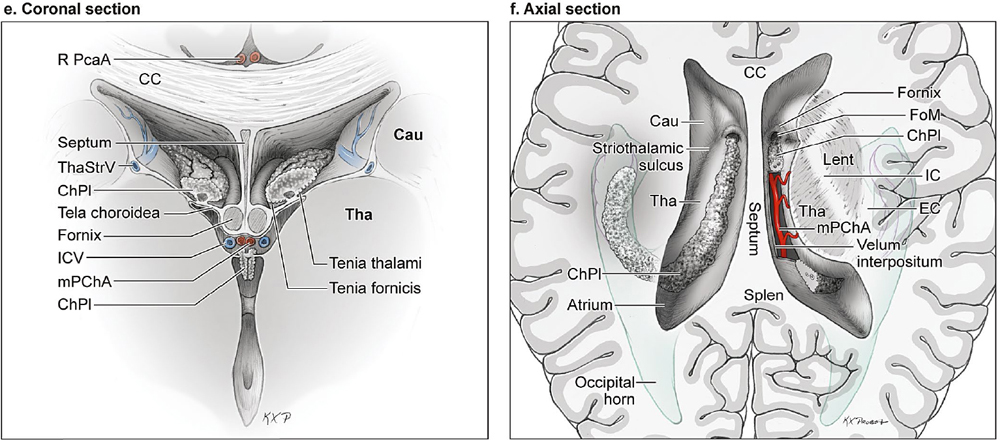
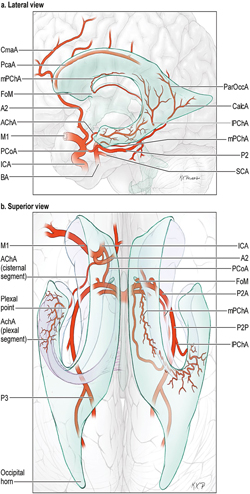
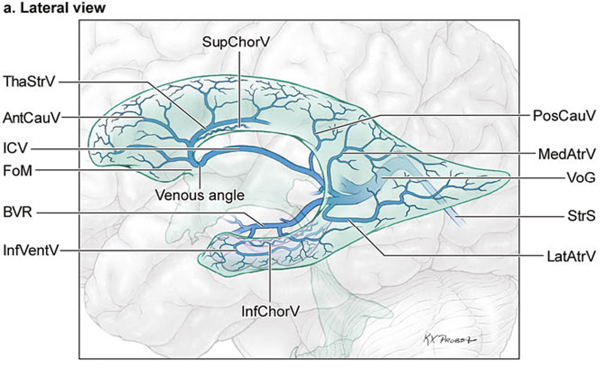
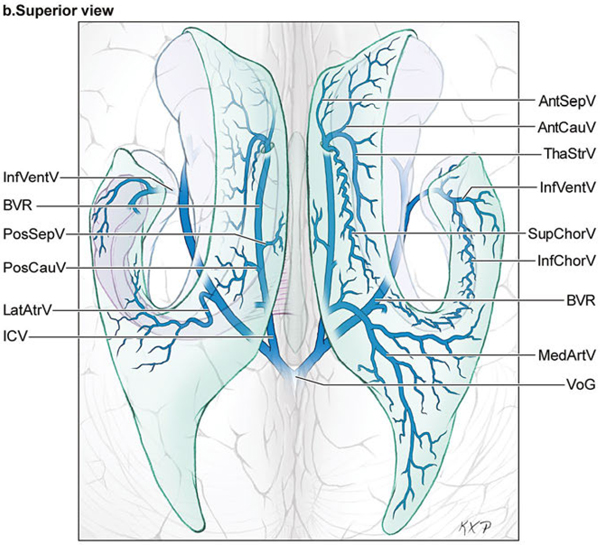
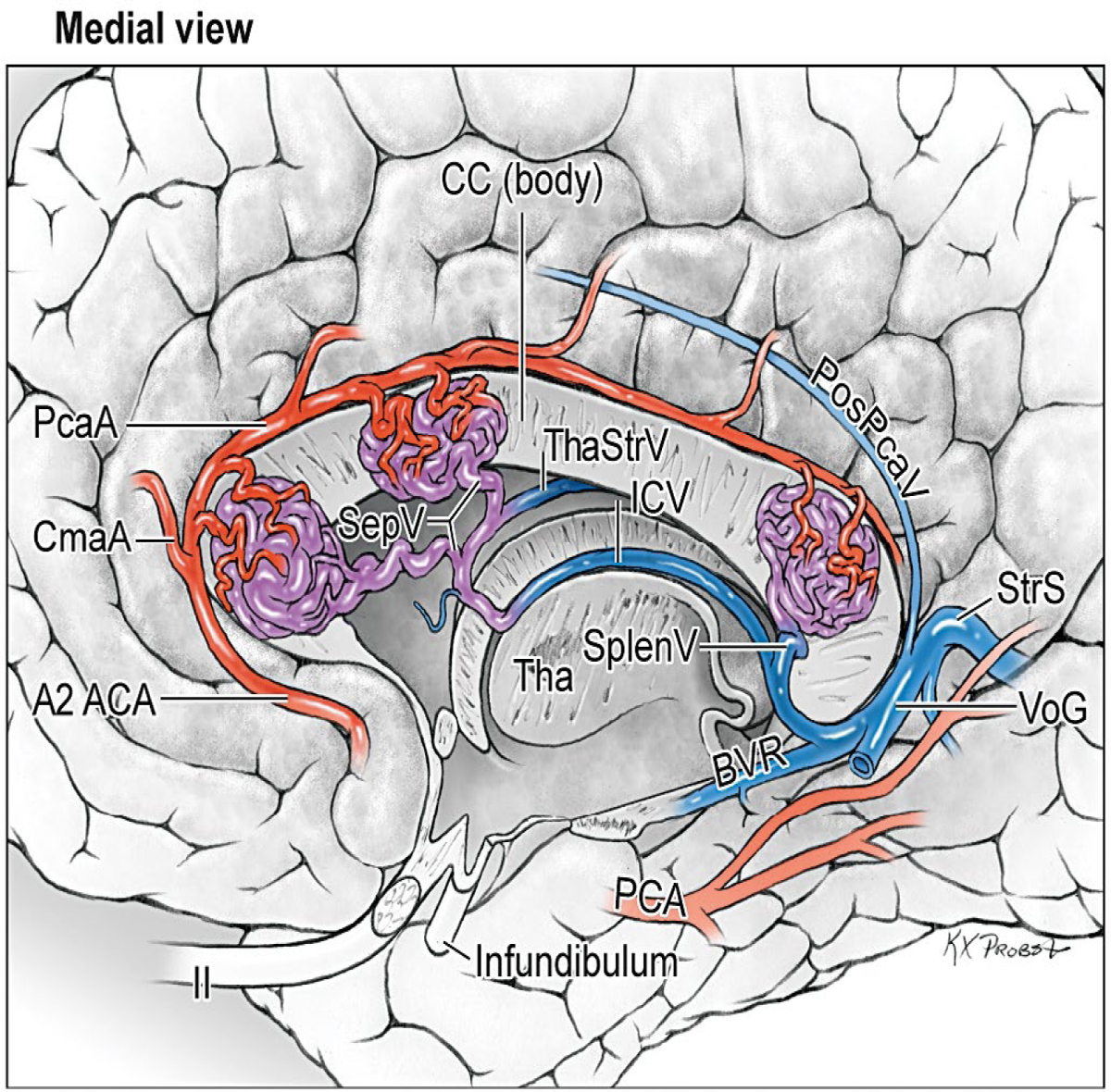
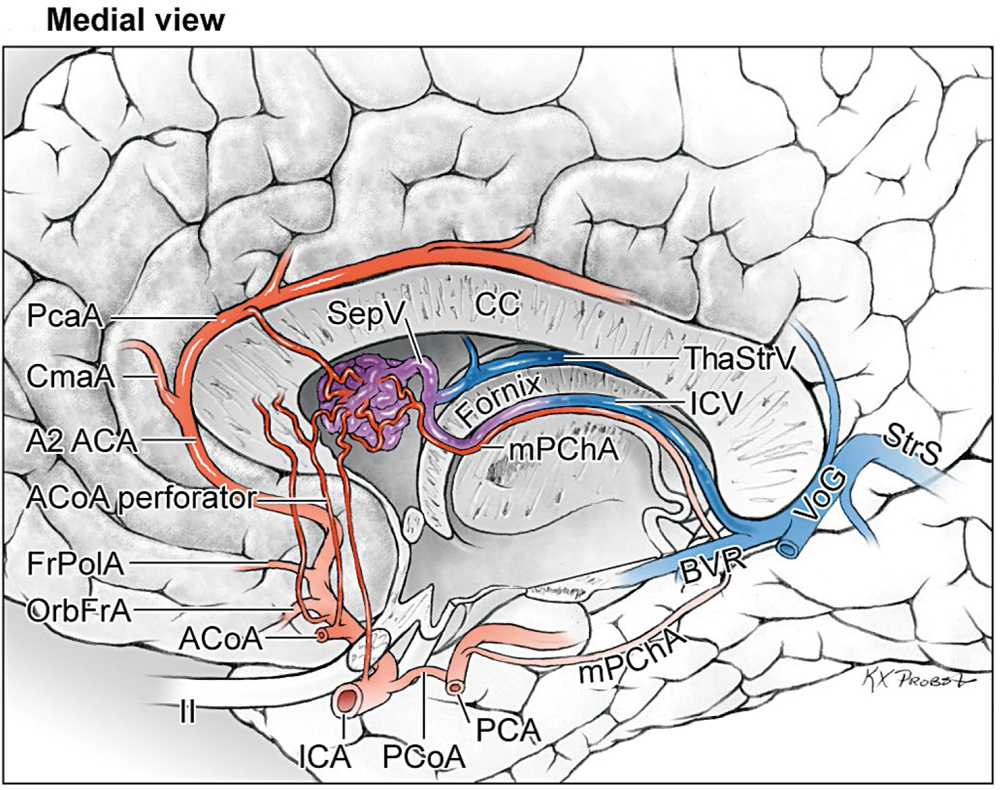
14 Ventricular and Periventricular Arteriovenous Malformations The cerebrum separates neatly into frontal, temporal, and parieto-occipital lobes, and arteriovenous malformations (AVMs) located in the cerebral hemispheres are based on their cortical surfaces. The ventricles and deep structures of the central core lie at the center of the cerebrum and lack cortical surfaces, but have ependymal surfaces on which ventricular and periventricular AVMs are based. Although the central core is composed of neural structures (basal ganglia, thalamus, internal, external, and extreme capsules, to be covered in the following chapter), the ventricles are cavities whose contents are simply cerebrospinal fluid (CSF) and choroid plexus. Ventricular and periventricular AVMs reside in the choroid plexus and some of the structures that make up the ventricle’s walls. The lateral ventricle is composed of five parts: frontal horn, body, atrium, occipital horn, and temporal horn (Fig. 14.1). Choroid plexus is present only in the body, atrium, and temporal horn; its absence in the frontal and occipital horns accounts for the absence of AVMs there. Choroid plexus in the body of the lateral ventricle and the roof of the third ventricle are close and continuous, running through each foramen of Monro (FoM), turning around 180 degrees to hang as parallel strands into the third ventricle, and running to the suprapineal recess posteriorly. The thalamus lies at the center of the cerebrum and the lateral ventricle wraps around it in a “C” shape. Three other C-shaped structures wrap around the thalamus superiorly, posteriorly, and inferiorly in the wall of the lateral ventricle: choroid plexus, fornix, and caudate nucleus. The fornix is composed of hippocampomammillary tract fibers that arise as fimbria on the ventricular surface of the hippocampal formation in temporal horn. They form the crus of the fornix, which courses posteriorly, wraps around the pulvinar of the thalamus, and joins the opposite crus in the body of the lateral ventricle to become the body of fornix. The fornix continues forward along the superomedial thalami in the medial wall of the lateral ventricular body, splits anteriorly into two columns, and then dives into the anterior margin of the FoM to end in the mammillary bodies. The caudate nucleus forms the lateral wall of the lateral ventricle with its head bulging into the ventricle at the frontal horn and body, its body forming part of the lateral wall at the atrium, and its tail curving into the roof of the temporal horn. Fig. 14.1 Microsurgical anatomy of the ventricles and periventricular structures. Five parts of the lateral ventricle (frontal horn, ventricular body, atrium, temporal horn, and occipital horn) and five C-shaped structures wrapping around the thalamus (lateral ventricle, caudate nucleus, fornix, choroid plexus, and corpus callosum) are shown in (a) anterior, (b) superior, and (c,d) lateral views. Microsurgical anatomy of the ventricular body and choroidal fissure, (e) coronal and (f) axial cross-sectional views, with the tela choroidea cut and a portion of the choroid plexus removed from the right ventricular body to show the mPChA in the velum interpositum. The corpus callosum forms the largest part of the ventricular walls, and is made up of the rostrum and genu anteriorly, the body centrally, and the splenium posteriorly. The rostrum forms the floor of the frontal horn; the genu and body form the roof; and the splenium forms the medial wall of the atrium. The septum pellucidum separates the lateral ventricles in the midline. It is composed of two laminae that are normally fused (but may also be separated as a cavum septum pellucidum) and attach to the genu and rostrum in the frontal horn and to the body and fornix in the ventricular body. The choroidal fissure is a narrow cleft between the fornix and thalamus where the choroid plexus attaches in the ventricular body, atrium, and temporal horn (Fig. 14.1). The choroidal fissure extends in a C-shaped arc from the FoM, around the thalamic surfaces, to the inferior choroidal point behind the head of hippocampus. The choroidal fissure is divided into the ventricular body and atrial and temporal parts. The tela choroidea is the clear membrane in which the choroid plexus arises, and the tela choroidea attaches to neural structures via small ridges called teniae: tenia thalami, tenia fornicis, and tenia fimbriae. The temporal part of the choroidal fissure is in the temporal horn between the fimbria of the fornix and thalamus. The body portion of the choroidal fissure opens into the velum interpositum and roof of third ventricle. The third ventricle is a narrow, funnel-shaped, midline cavity located below the body of the lateral ventricles between the two halves of thalamus and hypothalamus. Third ventricle communicates with each lateral ventricle through the FoM and with the fourth ventricle through the aqueduct of Sylvius. The roof of the third ventricle is composed of four layers: the fornix, two membranes of the tela choroidea, and a layer of blood vessels between the tela [the medial posterior choroidal artery (mPChA) and the internal cerebral vein (ICV)]. The velum interpositum is the space between the two layers of the tela in the roof of the third ventricle. Veins of the body and frontal horn converge on the ICV in the velum interpositum just behind the FoM. The confluence of arteries and veins traveling in the velum interpositum make this is an important region for ventricular AVMs. The ependyma and choroid plexus of the lateral and third ventricles are supplied by the anterior (AChA) and posterior choroidal arteries (mPChA and lateral posterior choroidal artery [lPChA]) (Fig. 14.2). The AChA arises from lateral supraclinoid internal carotid artery (ICA) as its last branch before its terminal bifurcation. Its initial “cisternal” segment has a characteristic posteromedial course, which on anteroposterior angiography is seen medial to the ICA. It passes below and medial to the optic tract to reach the lateral aspect of the cerebral peduncle and the crural cistern, where it runs between the cerebral peduncle and uncus. It climbs superiorly on the uncus to reach the choroidal fissure of the temporal lobe. This cisternal segment is bisected by the lateral geniculate body. The AChA’s intraventricular “plexal” segment begins at the plexal point where it pierces the choroidal fissure, enters the temporal horn, and supplies the choroid plexus. It continues posteriorly along the medial border of the choroidal plexus in close relation to and anastomosing frequently with branches of the lPChA. Critical branches from the cisternal segment supply the optic tract, the middle third of the cerebral peduncle, the posterior limb of the internal capsule, the lateral geniculate body, the globus pallidus, and the origin of optic radiations, whereas branches from the plexal segment are usually non-eloquent. Fig. 14.2 Microsurgical anatomy of the ventricular arteries. The ependyma and choroid plexus of the lateral and third ventricles are supplied by choroidal arteries (AChA, mPChA, and lPChA), (a) lateral and (b) superior views. The AChA has cisternal and plexal segments divided by the plexal point, where it traverses the choroidal fissure of the temporal horn. The mPChA has a long course from the P2 PCA to the atrium, encircling the midbrain, coursing through the velum interpositum, and running along the lateral ventricular floor, with two complete reversals of direction along the way. The lPChA has a straight course from the distal P2 PCA to the atrium. The lPChA originates from the distal P2 segment (ambient segment, P2P), courses laterally through the ambient cistern, and enters the choroidal fissure of the lateral ventricle at the posterior part of the temporal horn and atrium. There are multiple lPChAs (four on average) that anastomose with the mPChA as its runs posteriorly toward the atrium and with the AChA as it runs posteriorly in the temporal horn. The lPChA sends branches to the cerebral peduncle, posterior commissure, fornix (crura and body), lateral geniculate body, pulvinar, dorsomedial thalamic nucleus, and body of caudate nucleus. Corpus callosum is supplied by the anterior cerebral artery (ACA) and pericallosal artery (PcaA). Short callosal arteries from the PcaA penetrate the genu and body of the corpus callosum to supply these structures and the septum pellucidum, columns of fornix, and anterior commissure. Long callosal arteries run parallel to the PcaA and also supply the corpus callosum. Splenium is supplied by SplenA, which originates variously from ParOccA, CalcA, PosTempA, mPChA, or lPChA. SplenA anastomoses with the PcaA to complete the “limbic loop,” a collateral circuit around the corpus callosum that connects the ACA and PCA circulations. Ventricular veins on the lateral ventricular walls provide landmarks during intraventricular surgery that are often more orienting than the arteries, which are smaller, fewer in number, and buried below tufts of choroid plexus. Veins are easily visible in the ependyma (Fig. 14.3) and with AVMs, and are often dilated and tortuous. Anterior and posterior septal veins (SepV) drain the medial frontal horn and course to the FoM where they terminate in the ICV. Anterior and posterior caudate veins (CauV) drain the lateral frontal horn and terminate in the thalamostriate vein (ThaStrV) near the FoM. ThaStrV courses in the striothalamic sulcus between the caudate nucleus and thalamus. It makes a hairpin turn at the posterior margin of the FoM to enter the velum interpositum and join the ICV, a turn known angiographically as the “venous angle” on lateral views in the venous phase. The ICVs are paired, parallel veins running in the velum interpositum from the FoM to the vein of Galen (VoG). The two ICVs, two basal veins of Rosenthal (BVRs), and the precentral cerebellar vein converge on the VoG. Medial atrial veins (AtrV) drain the medial wall of the atrium, passing through the fornix or choroidal fissure to the ICV in the velum interpositum. The lateral AtrV drains the lateral wall of the atrium and passes through the choroidal fissure to reach the BVR in the ambient or quadrigeminal cisterns. The medial and lateral AtrV can join near the choroidal fissure to form a common trunk, the common atrial vein. The superior choroidal vein (ChorV), the largest of the choroidal veins, runs in the choroid plexus in the body of the lateral ventricle and terminates in ThaStrV in the FoM. The inferior ChorV runs in the temporal horn and terminates in the inferior ventricular vein or the amygdaloid vein, which then exits the choroidal fissure to join the BVR. The superior and inferior ChorVs anastomose in the glomus of the choroid plexus in the atrium. Fig. 14.3 Microsurgical anatomy of ventricular veins and deep venous system. (a) Veins of the lateral ventricle, lateral and (b) superior views. Ventricular and periventricular AVM subtypes include callosal, ventricular body, atrial, and temporal horn AVMs. Callosal AVMs are located in the roof of the lateral ventricles (Fig. 14.4). This subtype was the most common, accounting for more than half of ventricular/periventricular AVMs. The classic AVM is entirely in the corpus callosum, but it can extend superiorly to involve some cingulate gyrus or inferiorly into the septum or lateral ventricle. Callosal AVM locations range from rostrum to splenium with important anatomic and surgical differences, but they are considered collectively because of their relative rarity (approximately 4% of all brain AVMs). They are supplied by the ACA and PcaA, which give rise bilaterally to short and long callosal arteries on their inferior and medial surfaces that descend into the corpus callosum and AVM. These feeding arteries are tiny in the absence of callosal AVMs but enlarge in their presence. Normal branches to the cingulate gyrus and medial frontal lobe can be differentiated from AVM feeders by their small size and their origin from PcaA’s lateral surface. The PcaA is typically a transit artery that supplies the medial homunculus for the motor and sensory cortex distal to the AVM, and therefore must be preserved by skeletonization. Although anterior callosal AVMs in the genu and body are supplied by the PcaA, posterior callosal AVMs are also supplied by SplenA from the PCA. Callosal AVMs that extend laterally into the frontal white matter can get additional supply from the medial lenticulostriate artery (mLSA) or lateral lenticulostriate artery (lLSA). Although they are included with ventricular AVMs, callosal AVMs do not involve the choroid plexus and are not supplied by the choroidal arteries. Callosal AVMs drain deep into ventricular veins (SepV, CauV, ThaStrV, and ICV). Splenial AVMs also drain deep to the VoG, either via SplenV that remains extraventricular or through MedAtrV that descends to the ICV distal to the FoM. Callosal AVMs are not strictly eloquent, but the splenium has been implicated in calculation and visuospatial processing. Fig. 14.4 The callosal AVM, medial view. Callosal AVMs are located in the corpus callosum and range from the rostrum to the splenium; three different callosal AVMs are shown in the genu, body, and splenium. They are supplied by long and short callosal arteries branching from the ACA and PcaA, and drain deeply to ventricular veins (SepV, CauV, and ThaStrV). The ventricular body AVM is a midline AVM located in the center of the lateral ventricular system at the complex intersection of the thalamus, caudate nucleus, fornix, septum, velum interpositum, choroid plexus, and choroidal fissure (Fig. 14.5). Ventricular body AVMs include those in the septum, body of fornix, velum interpositum, and choroid plexus of the ventricular body or third ventricle. Ventricular body AVMs do not include those in the lateral wall, lateral floor, or roof of the ventricular body. The caudate nucleus forms the lateral wall, and AVMs here are included in the basal ganglial subtype of deep AVMs; the thalamus forms the lateral floor, and AVMs here are included in the thalamic subtype of deep AVMs; and the corpus callosum forms the roof of the ventricular body and AVMs here are included in the callosal subtype. Ventricular body AVMs are uncommon, accounting for only 12% of ventricular/periventricular AVMs. They are challenging because they are deeper than callosal AVMs and accessible only through a small opening in the corpus callosum and choroidal fissure. The mPChA runs through the center of the region and is the major feeder to these AVMs. The lPChA supply is usually minimal. The PcaA contributes to AVMs in the septum. The mLSA and anterior communicating artery (ACoA) perforators can supply septal AVMs anterior to the FoM. The ICV also runs through the center of the region and drains these AVMs. The fornix is intimately related to these AVMs, forming the medial border of the choroidal fissure and also descending in the anterior aspect of the FoM. Even though the ventricular body AVM is surrounded by CSF rather than parenchyma, forniceal manipulation can cause debilitating memory and cognitive impairment, making it an eloquent AVM that requires careful handling. The atrial AVM resides in the atrium, posterior and lateral to the ventricular body (Fig. 14.6). Callosal and ventricular body AVMs are midline, whereas atrial and temporal horn AVMs have laterality. The C-shaped structures that wrap around the thalamus (i.e., caudate nucleus, crus of fornix, and choroid plexus) deviate laterally as they course around its posterior surface toward the temporal horn, and atrial AVMs are in the middle zone between the ventricular body and temporal horn. They were the second most common ventricular/periventricular subtype and accounted for around 20%. Atrial AVMs are supplied by the lPChA, which enters the ventricle at the atrial portion of the choroidal fissure. Collateral connections within the choroid plexus enable the AChA to supply these AVMs from below via the temporal horn’s plexus, and the mPChA to supply them from anteriorly via the ventricular body’s plexus. Atrial AVMs drain through the medial and lateral AtrVs that course through the choroidal fissure of the atrium and connect to the ICV and BVR, respectively. Atrial AVMs sit on the pulvinar surface, which has no eloquence, but the crus of the fornix lies medial to the choroid plexus. Atrial AVMs differ from thalamic AVMs that may protrude into the atrium, but are deeply embedded in the thalamus, extend close to the posterior limb of the internal capsule, are supplied by thalamoperforators rather than choroidal arteries, and are exquisitely eloquent. Fig. 14.5 The ventricular body AVM, medial view. Ventricular body AVMs include those in the septum, body of fornix, velum interpositum, and choroid plexus of the ventricular body and the roof of third ventricle. The mPChA is a consistent supplier to these AVMs, with additional supply from the PcaA, ACoA perforators, mLSA, and lPChA, depending on location. Venous drainage collects in the internal cerebral vein.
 Microsurgical Anatomy
Microsurgical Anatomy
Brain
Arteries
Veins
 Four Ventricular AVM Subtypes
Four Ventricular AVM Subtypes
The Callosal AVM
The Ventricular Atrium AVM
The Ventricular Body AVM
Related posts:
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Neupsy Key
Fastest Neupsy Insight Engine


