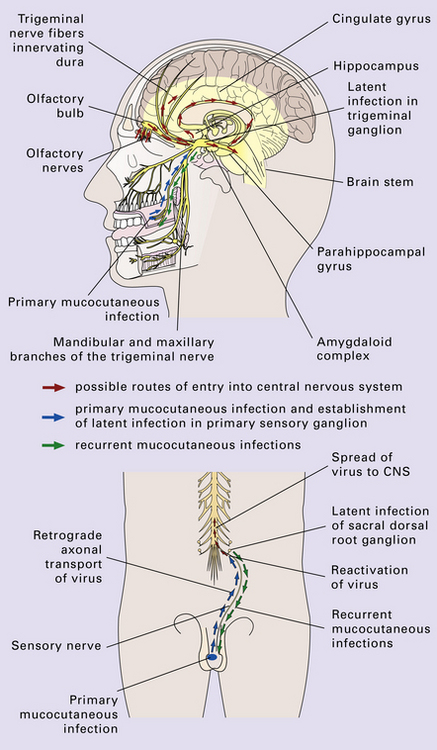
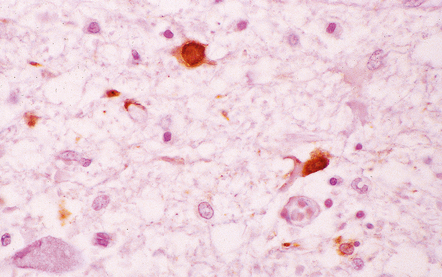
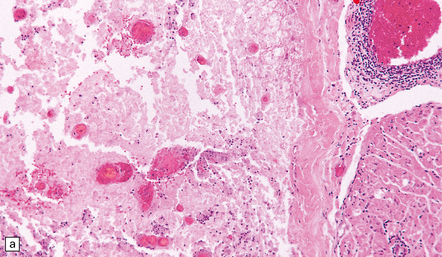
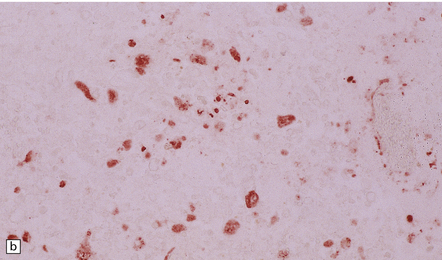
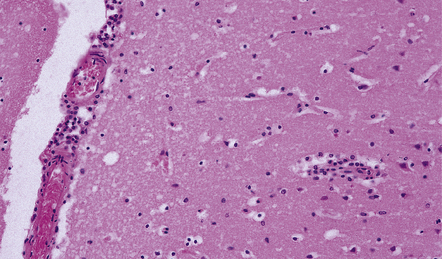
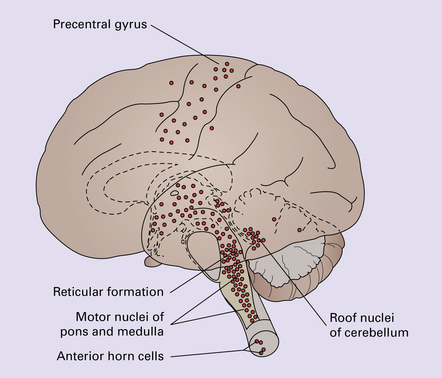
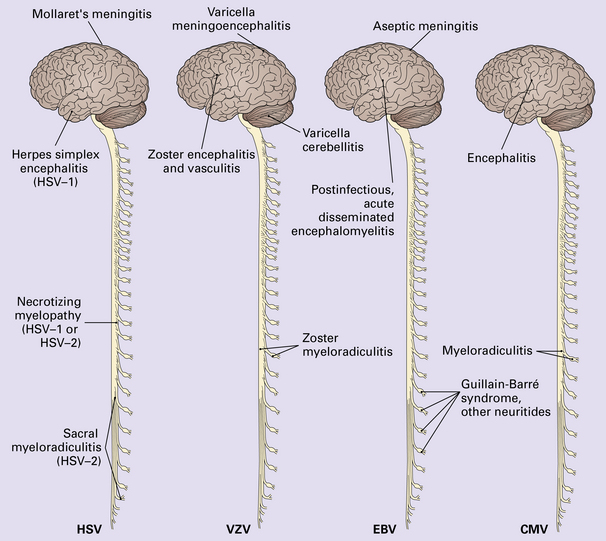
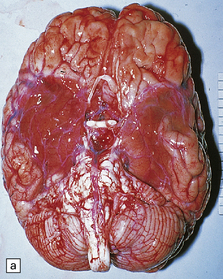
12 Classifications of viral infections of the CNS such as those shown in Table 12.1 are of help in making an accurate diagnosis. In practice, a combination of approaches is generally used to classify and diagnose these disorders. Table 12.1 Viral infections of the central nervous system These may be classified according to: Course of disease Acute Subacute or chronic Most severely involved/target tissue Meningitis Poliomyelitis, polioencephalomyelitis Viral leukoencephalopathy Panencephalitis Patient age group Fetal Neonatal Child Adult Patient immune status Etiologic agent This is a benign, usually short-lived, syndrome of meningeal inflammation that is not attributable to any of the common bacterial pathogens. A wide range of viruses can cause aseptic meningitis (Table 12.2); much less common causes include some bacteria (e.g. in syphilis and Lyme disease) and other microorganisms, non-infective inflammatory disorders (e.g. Behçet’s disease), tumors (e.g. epidermoid and dermoid cysts), and drugs (e.g. ibuprofen). One form of recurrent aseptic meningitis (Mollaret’s) that was previously regarded as non-infective has been linked to infection with herpes simplex virus (HSV), especially HSV-2. Table 12.2 Common viral causes of aseptic meningitis Echovirus Coxsackie B Coxsackie A Herpes simplex virus (HSV)-2 Mumps Human immunodeficiency virus (HIV) Lymphochoriomeningitis virus Arboviruses Measles Parainfluenza virus Adenovirus Since aseptic meningitis is by definition benign, reports of the neuropathologic findings are uncommon. Occasionally, patients with aseptic meningitis die as a result of a concurrent systemic illness (e.g. viral myocarditis). Histologic examination of the CNS then reveals a scanty infiltrate of lymphocytes in the meninges, in the perivascular space surrounding some of the superficial cortical blood vessels (Fig. 12.1), and in the choroid plexus. The extent of histologic involvement almost always exceeds that predicted by the clinical manifestations. The distribution of lesions is variable. The spinal gray matter is usually involved, particularly the anterior horn cells. The disease also shows a predilection for the motor nuclei in the pons and medulla, the reticular formation, and deep cerebellar nuclei (Fig. 12.2). Apart from the precentral gyrus, the cerebral cortex is usually spared. There is intense inflammation in the leptomeninges and affected gray matter (Fig. 12.3). Neutrophils are found initially, but lymphocytes soon predominate. Lymphocytic cuffing of blood vessels is a conspicuous feature. 12.3 Poliomyelitis. The histologic hallmark of the viral infection of neurons is neuronophagia (aggregation of microglia and macrophages around dead neurons, Fig. 12.4). Clusters of microglia (microglial nodules) mark the sites of destroyed neurons for several weeks after their resorption. 12.4 Neuronophagia and microglial nodule formation in poliovirus infection. Examination of the affected regions shows an obvious loss of motor neurons (Fig. 12.5) and atrophy and fibrosis of anterior nerve roots. There may be scanty residual inflammation. These changes apart, the parenchyma of the affected spinal cord or brain stem is usually remarkably well preserved. 12.5 Chronic phase of poliomyelitis. The differential diagnosis of polioencephalomyelitis includes infection due to these and other arboviruses (see below), rabies (see below) and non-infective encephalitides, particularly Rasmussen’s encephalitis (see Chapters 7 and 13) and paraneoplastic (autoimmune) encephalitis (see Chapter 47). The appearances differ from those in older children and adults. Although the CNS lesions consist of infiltrates of lymphocytes, macrophages and microglia and are usually centered mainly on the gray matter of the brain stem and spinal cord, the white matter is often affected, the lesions may be necrotizing or hemorrhagic, and foci of cerebellar and cerebral inflammation are common (Fig. 12.6). 12.6 Neonatal enteroviral infections often cause destructive infection of multiple organs, and although the CNS disease is predominantly a polioencephalomyelitis, the white matter may also be involved. The herpesviruses are relatively large, enveloped, double-stranded DNA viruses. The group includes several that are human pathogens and can cause CNS disease (Fig. 12.7), including herpes simplex virus type 1, herpes simplex virus type 2, Epstein–Barr virus, cytomegalovirus, and human herpesvirus 6. The simian herpesvirus, B virus, can also infect humans and cause CNS disease. When these viruses invade the CNS they tend to cause necrotizing destruction of both gray and white matter (i.e. panencephalitis or panmyelitis). CLASSICAL HERPES SIMPLEX ENCEPHALITIS (HSE) Most cases show obvious congestion and hemorrhagic necrosis involving the temporal lobes (Fig. 12.9) and, to a greater or lesser extent, the insulae, cingulate gyri, and posterior orbital frontal cortex (Fig. 12.9). The lesions are often somewhat asymmetric. Occasionally, in very early disease, the brain may appear macroscopically normal. In contrast, in patients dying some weeks after the onset of disease the liquefactive necrosis in these regions will have progressed to cavitation and atrophy. The earliest lesions contain relatively scanty parenchymal inflammation, although there are moderate numbers of lymphocytes and macrophages in the overlying leptomeninges (Fig. 12.10). The lesions extend from the pial surface through the cerebral cortex and into the white matter. The affected neurons, glia, and endothelial cells tend to have slightly hypereosinophilic cytoplasm. Many of the nuclei are pyknotic or disintegrating; others contain homogeneous eosinophilic inclusions (Fig. 12.10), some surrounded by an irregular rim of condensed marginated chromatin. Clumps of eosinophilic inclusion material may also be visible in the cytoplasm. Inclusions are usually best seen in cells towards the edge of lesions. 12.10 Early HSE. Most lesions are usually at a more advanced stage, containing sheets of necrotic cells, foci of hemorrhage, and an intense perivascular and interstitial infiltrate of lymphocytes and macrophages (Fig. 12.11). There may be neuronophagia and, later, microglial nodules. Nuclear inclusions are sparse at this stage. 12.11 Necrotizing inflammation in HSE. Herpesvirus nucleocapsid particles are approximately 100 nm in diameter and may be seen within the nuclei of infected cells by electron microscopy (Fig. 12.12). Viral antigen is readily demonstrable by immunohistochemistry (Fig. 12.13) for up to approximately 3 weeks after the onset of encephalitis, and viral DNA can be detected in frozen or paraffin sections by in situ hybridization (Fig. 12.14) or polymerase chain reaction (PCR) amplification with suitable primers. 12.12 Ultrastructural appearance of HSV. 12.13 Immunohistochemical demonstration of HSV antigen, some of which is clearly within neurons (arrows). Primary mucocutaneous infection In most patients, initial infection by HSV-1 involves the mucocutaneous border of the lips or the oropharyngeal mucosa. Establishment of latency in the trigeminal ganglion After local replication the virus is conveyed by retrograde axonal transport along sensory fibers to the trigeminal ganglion, where, after further replication, latent infection is established (Fig. 12.8). (In contrast, HSV-2 causes genital herpes infection and establishes latency in the sacral dorsal root ganglia.) HSV-1 genome and latency-associated transcripts (the only viral mRNAs produced during latent infection) can be detected in the trigeminal ganglia in 50–75% of adults. Involvement of the olfactory pathway HSV-1 DNA has been detected in the olfactory bulbs in approximately 15% of adults, suggesting that retrograde transport of the virus along the olfactory nerve fibers may occur after primary nasopharyngeal infection. It is not known whether the virus in the olfactory bulbs is susceptible to reactivation. In experimental studies, attempts at reactivation from central nervous tissue have been unsuccessful. The mechanism of entry of HSV-1 into the CNS to cause HSE has been much debated. Proposals include: In long-term survivors of untreated or unsuccessfully treated herpes encephalitis, affected parts of the brain are shrunken and cavitated and show yellow-brown discoloration (Fig. 12.15). 12.15 ‘Burnt out’ HSE. The normal gray and white matter is replaced by cavitated glial scar tissue (Fig. 12.15). Occasional clusters of lymphocytes are still seen in the meninges and brain parenchyma (Fig. 12.15). In occasional patients, HSE predominantly involves the brain stem or follows a more subacute clinical course than usual (Fig. 12.16). Although the likelihood of developing HSE is probably not increased by immunosuppression, immunosuppression predisposes to atypical forms of the disease. Very rarely, children who have experienced an otherwise typical attack of acute herpes encephalitis develop focal or multifocal chronic granulomatous encephalitis, sometimes after an intervening symptom-free period of months or years. Histology reveals a patchy cortical and leptomeningeal infiltrate of chronic inflammatory cells and scattered, well-circumscribed granulomas that contain epithelioid macrophages and giant cells, with surrounding lymphocytes, macrophages, and plasma cells. Foci of necrosis and mineralization may be prominent. In some patients, HSV DNA or antigen is demonstrable by PCR or immunohistochemistry (Fig. 12.17). 12.17 Chronic granulomatous herpes simplex encephalitis. Both HSV-1 and HSV-2 can cause this rare disorder. Histology shows extensive necrosis and inflammation involving the spinal gray and white matter (Fig. 12.18). Herpesvirus is usually demonstrable within the cord by immunohistochemistry (Fig. 12.18).
Acute viral infections
ASEPTIC MENINGITIS
MACROSCOPIC AND MICROSCOPIC APPEARANCES
POLIOMYELITIS
MICROSCOPIC APPEARANCES
Acute phase
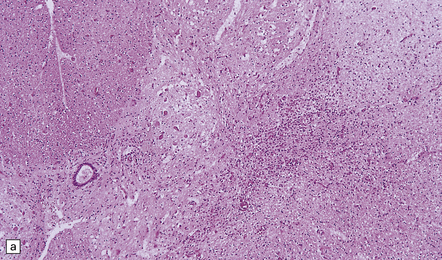
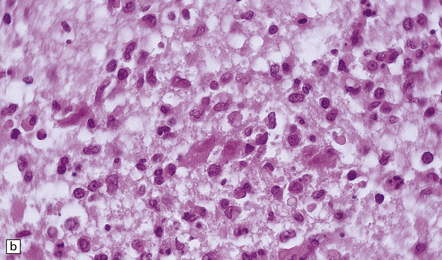
(a) and (b) show mixed inflammatory infiltrate in the spinal gray matter in coxsackievirus poliomyelitis. (Courtesy of Dr David Hilton, Derriford Hospital, Plymouth, UK.)
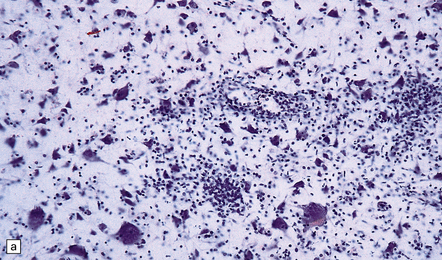
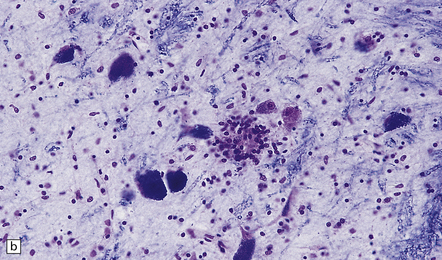
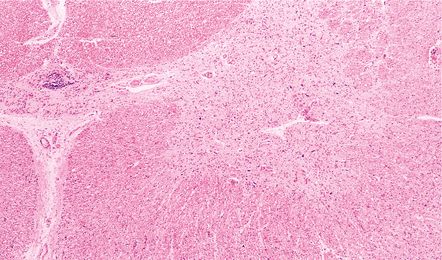
(a) Florid brain stem inflammation and neuronophagia in poliovirus poliomyelitis. (b) Microglial nodule in the brain stem in coxsackievirus polioencephalomyelitis.
MACROSCOPIC AND MICROSCOPIC APPEARANCES
Chronic phase
Marked depletion of anterior horn cells from the spinal cord of a long-term survivor of poliomyelitis.
 Most patients experience no more than a minor nonspecific illness at the time of the primary viremia, 1–5 days after exposure to the virus. Symptoms include: gastrointestinal upset, mild pyrexia, headache, and general malaise.
Most patients experience no more than a minor nonspecific illness at the time of the primary viremia, 1–5 days after exposure to the virus. Symptoms include: gastrointestinal upset, mild pyrexia, headache, and general malaise.
 Patients may experience muscle pain or stiffness before the development of paralysis.
Patients may experience muscle pain or stiffness before the development of paralysis.
 CAUSES OF POLIOMYELITIS AND POLIOENC EPHALOMYELITIS
CAUSES OF POLIOMYELITIS AND POLIOENC EPHALOMYELITIS
NEONATAL ENTEROVIRAL ENCEPHALITIS
MICROSCOPIC APPEARANCES
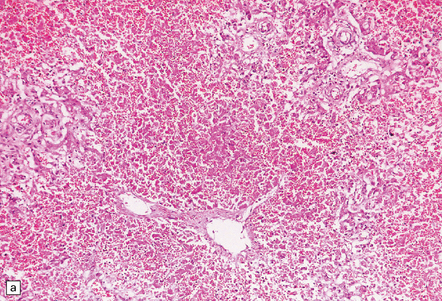
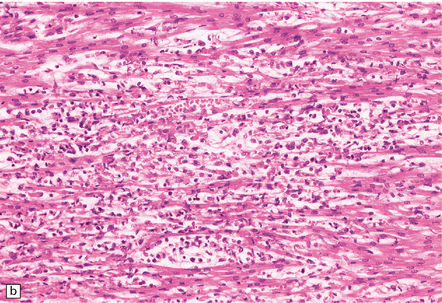
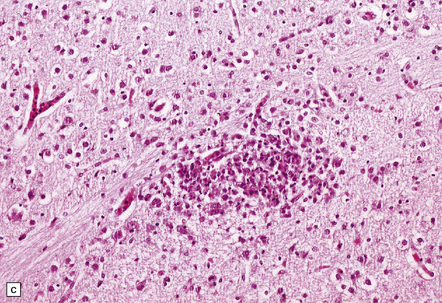
(a) Neonatal coxsackievirus B3 infection of the liver has caused extensive panlobular necrosis. (b) There is also myocarditis, with extensive infiltration by mononuclear inflammatory cells. (c) A cluster of lymphocytes, macrophages, and microglia is present in the tegmentum of the pons. (d) Inflammatory cell infiltrates are present both in the white matter (arrow) and the superficial part of the granule cell layer (arrowheads) in the cerebellum.
HERPESVIRUS INFECTIONS
HERPES SIMPLEX VIRUS INFECTION
MACROSCOPIC APPEARANCES
Acute phase
MICROSCOPIC APPEARANCES
Acute phase
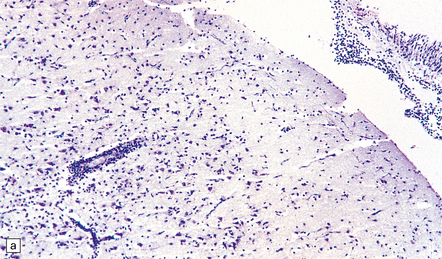
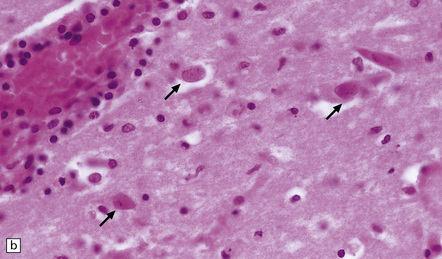
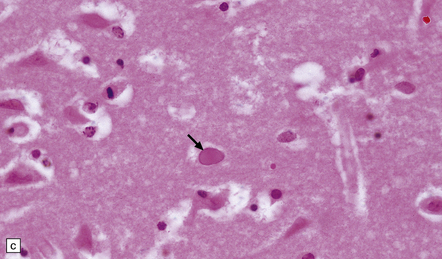
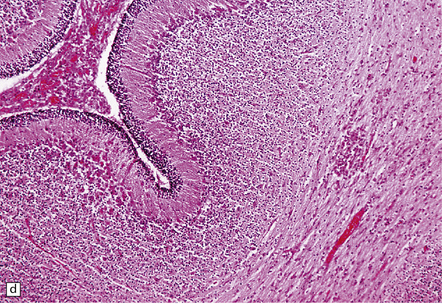
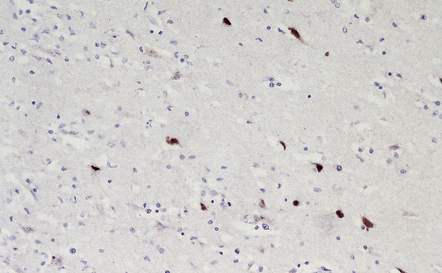
(a) Meningeal, perivascular, and scanty parenchymal inflammation. (b) This section shows scanty perivascular inflammation and scattered cells containing viral inclusions (arrows). (c) Towards the edge of the affected region of the temporal lobe, intranuclear viral inclusions (arrow) are visible.
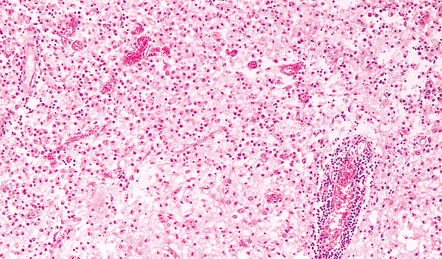
Sheets of foamy macrophages and perivascular cuffing by lymphocytes in a case of HSE.
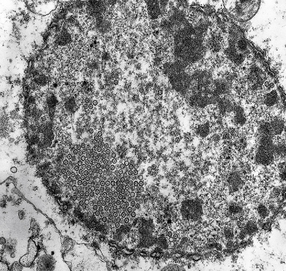
Electron micrograph showing intranuclear herpesvirus nucleocapsid particles.
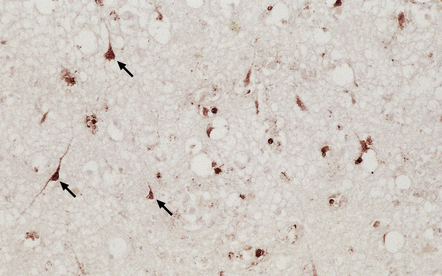
MACROSCOPIC AND MICROSCOPIC APPEARANCES
Chronic phase
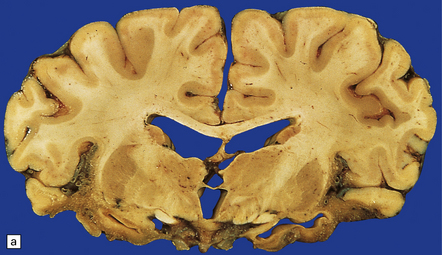
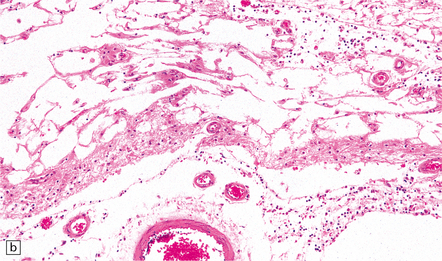
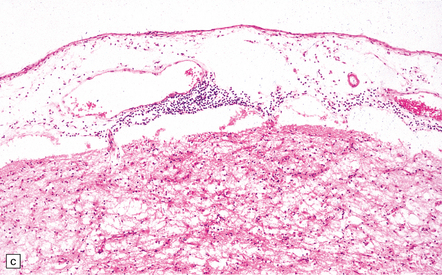
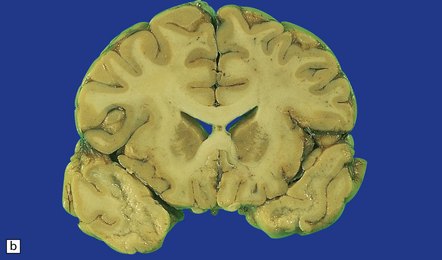
(a) Marked atrophy and yellow-brown discoloration affect the temporal lobes and insulae. (b) Cavitated temporal lobe in a long-term survivor of HSE. (c) Scattered aggregates of lymphocytes persist in the meninges or brain parenchyma for many years. Note also the cortical gliosis and microcystic change.
ATYPICAL HERPES SIMPLEX ENCEPHALITIS
CHRONIC GRANULOMATOUS HERPES SIMPLEX ENCEPHALITIS
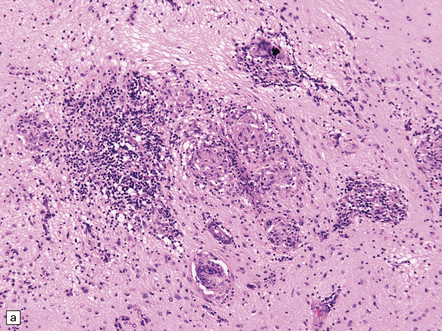
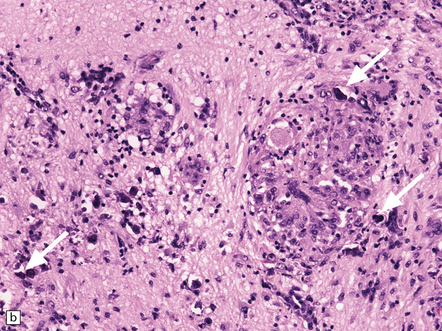
(a) Histology of partial temporal lobectomy specimen from a 10-year old boy with chronic granulomatous inflammation complicating herpes simplex virus encephalitis at 4.5 months of age reveals multiple granulomas, some including multinucleated giant cells, with a surrounding infiltrate of lymphocytes and macrophages. The arrow indicates a focus of mineralization. Postoperatively, HSV-1 DNA and elevated titers of HSV IgM antibodies were detected in the CSF. (b) Higher magnification view of the granulomas and foci of mineralization (arrows).
NECROTIZING MYELOPATHY
NEONATAL HSV ENCEPHALITIS
 PATHOGENESIS OF NEONATAL HSV ENCEPHALITIS
PATHOGENESIS OF NEONATAL HSV ENCEPHALITIS
 Most cases of neonatal HSV infection are due to HSV-2 and are acquired during delivery by contact between the fetus and infected maternal genital lesions. The disease can, however, be acquired from the mother in utero, even during early gestation, or by contact with infected secretions from the mother or another source during the postnatal period.
Most cases of neonatal HSV infection are due to HSV-2 and are acquired during delivery by contact between the fetus and infected maternal genital lesions. The disease can, however, be acquired from the mother in utero, even during early gestation, or by contact with infected secretions from the mother or another source during the postnatal period.
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Acute viral infections
 POLIOMYELITIS
POLIOMYELITIS




 POLIOVIRUS INFECTION
POLIOVIRUS INFECTION






 HERPES SIMPLEX ENCEPHALITIS
HERPES SIMPLEX ENCEPHALITIS


 HERPES SIMPLEX ENCEPHALITIS
HERPES SIMPLEX ENCEPHALITIS