FIGURE 11.1 The sulcus limitans divides the neural tube into the dorsal alar plate, which contains sensory neuroblasts, and the ventral basal plate, which contains motor neuroblasts. In the brainstem, the sulcus limitans separates the motor nuclei from the sensory nuclei.
As the brainstem develops, the expansion of the cavity of the fourth ventricle pushes the alar plate outward and downward. This causes the alar plate to retroflex so that it comes to lie lateral to, rather than dorsal to, the basal plate. The two plates are separated by the sulcus limitans (Figure 11.2). In the mature brainstem, the motor neurons derived from the basal plate lie medially, and the sensory neurons derived from the alar plate lie laterally. The neurons form cell columns, which are divided by anatomists into functional categories. The formal anatomical classification is somewhat arcane, seldom used by clinicians, but its conceptual framework is useful. The cell columns are divided into motor (efferent) and sensory (afferent) and into general and special, somatic and visceral cell types.
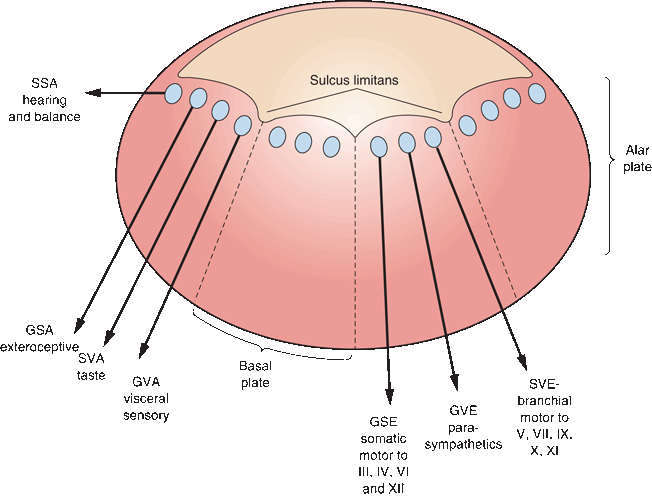
FIGURE 11.2 The cell columns of the brainstem with motor nuclei medial and sensory nuclei lateral. GSE, general somatic efferent; GVE, general visceral efferent; SVE, special visceral efferent; GVA, general visceral afferent; SVA, special visceral afferent; GSA, general somatic afferent; SSA, special somatic afferent.
Referring to Figure 11.2 and moving from medial to lateral, the first cell column is general somatic efferent (GSE), which contains somatic motor cells. The GSE cells innervate skeletal muscles, which are derived from myotomes. For the head and neck, these are the extraocular muscles and the tongue. The next cell column laterally is general visceral efferent (GVE). This contains visceral motor or autonomic (parasympathetic) neurons supplying smooth muscles and glands of the head and neck, and the thoracic and abdominal viscera as far as the splenic flexure of the colon. The GVE nuclei are the cranial portion of the craniosacral autonomic system, and they include the superior and inferior salivatory nuclei and the dorsal motor nucleus of the vagus.
Lateral to the GVE column is the special visceral efferent (SVE) column. Nuclei-innervating muscles of branchial (pharyngeal) arch origin were designated visceral because the gills in fish are derived from the embryonic branchial arches (Gr. branchia “gills”). Gills are viscera, primordial lungs, so branchial-arch-derived muscles in humans were considered visceral, and branchiomeric neurons were designated as SVE. In the course of ontogeny and phylogeny, the branchiomotor cell column drifted ventrally from a location just under the ventricular floor to a position in the tegmentum. The motor nuclei of CNs V and VII and the nucleus ambiguus lie about midway between the somatic motor nuclear column and the point of exit of their respective nerves. The nucleus ambiguus is obscured by the fibers of the reticular formation (RF), making it inconspicuous and difficult to identify. Because of the displacement of their nuclear columns, branchiomotor axons have a tendency to form internal loops, for example, the encirclement of the CN VI nucleus (GSE) by the axons leaving the CN VII nucleus (SVE). Somatic motor fibers exit the brainstem anteriorly; branchiomotor fibers exit laterally.
The sulcus limitans separates the most lateral motor cell column from the most medial sensory cell column. Most medial are the general visceral afferent—or visceral sensory—cell columns, which receive sensory input from the viscera. The special visceral afferent—or special sensory—cell column receives fibers subserving taste. The general somatic afferent column receives exteroceptive input (i.e., touch, pressure, pain, temperature, vibration, and proprioception) from the head and neck. The most lateral sensory column is for special somatic afferent functions subserving the special sensations (i.e., hearing and vestibular function).
External Anatomy
Selected major features of the external anatomy of the brainstem are shown in Figures 11.3 and 11.4. On the ventral surface, the rostral limit of the brainstem is demarcated by the optic tracts as they sweep around to reach the lateral geniculate bodies. Descending from beneath the optic tracts are the massive cerebral peduncles. The space between the peduncles is the interpeduncular fossa. At the upper margin of the interpeduncular fossa are the mammillary bodies. In its depths is the posterior perforated substance where paramedian perforating vessels from the basilar artery penetrate the upper brainstem and thalamus. CN III (oculomotor) emerges from the fossa and runs forward between the superior cerebellar and posterior cerebral arteries.
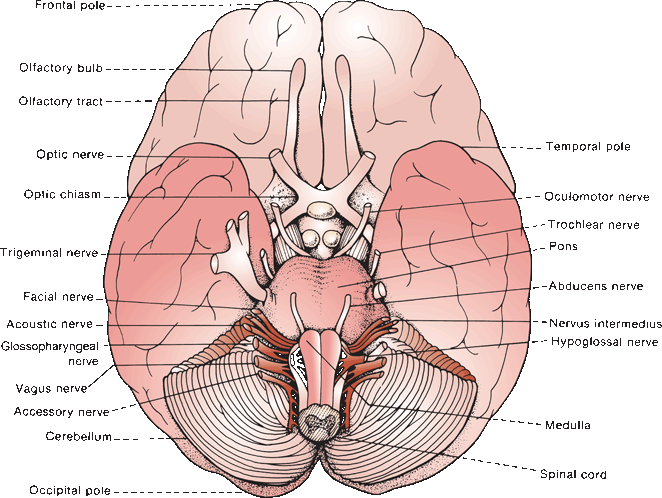
FIGURE 11.3 The base of the brain showing the sites of emergence of the cranial nerves.
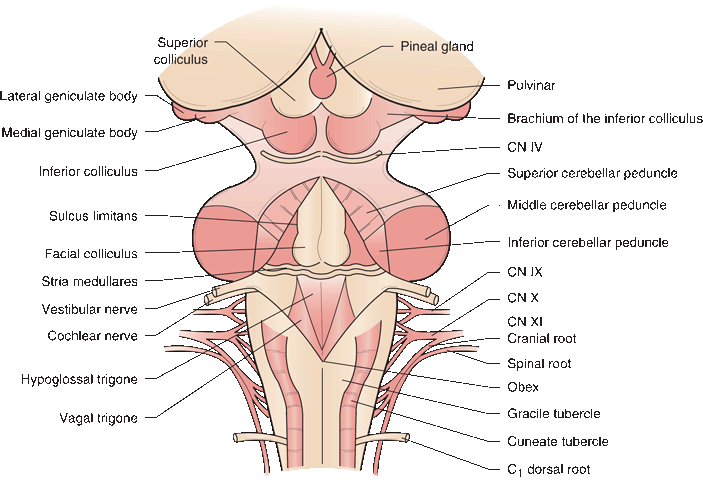
FIGURE 11.4 Dorsal view of the brainstem and rhomboid fossa.
At the caudal limit of the interpeduncular fossa is the junction between the midbrain and pons. The bulge of the anterior pons, due primarily to the underlying fibers of the middle cerebellar peduncle (MCP) (brachium pontis), bridges the space between the two cerebellar hemispheres and spans the gap between the midbrain and medulla (L. pons “bridge”). The root of CN V (trigeminal) is attached laterally at the level of the midpons. The furrow of the basilar artery, the basilar sulcus, grooves the pons from below to above. At the pontomedullary junction, from medial to lateral, CNs VI (abducens), VII (facial), and VIII (acoustic) exit. The nervus intermedius lies just lateral to the main root of the facial nerve. The vestibular division of CN VIII lies medial and slightly rostral to the cochlear division.
The medulla is 24 to 30 mm in length, and it extends from the pontomedullary junction and the striae medullares above to the lowermost roots of the hypoglossal nerve and the lowest plane of the pyramidal decussation—just rostral to the emergence of the highest rootlets of C1 at the level of the foramen magnum. Running down the anterior aspect of the medulla are the twin columns of the medullary pyramids, which contain the corticospinal tracts. Interlacing bundles of crossing fibers at the caudal-most extent of the medulla mark the decussation of the pyramids. Caudal to the decussation is the spinal cord. Just lateral to the pyramids in the upper medulla is the oval bulge of the olive, beneath which lies the inferior olivary nucleus. The CN XII (hypoglossal) filaments exit in the gutter between the pyramid and the olive. CNs IX (glossopharyngeal), X (vagus), and the cranial root of XI (accessory) exit in the retroolivary sulcus, in sequence from rostral to caudal.
Figure 11.4 shows the brainstem with the cerebellum removed and the fourth ventricle opened. The most rostral extent of the brainstem is marked by its junction with the pulvinar of the thalamus. The prominent mounds of the superior and inferior colliculi form the quadrigeminal plate. The pineal body extends caudally between the superior colliculi. The superior colliculus is connected to the lateral geniculate body by the brachium of the superior colliculus, and the inferior colliculus to the medial geniculate by its brachium. Just caudal to the inferior colliculus CN IV (trochlear) exits.
The fourth ventricular floor is rhomboid or diamond shaped and is called the rhomboid fossa. The superior cerebellar peduncle (SCP) forms the upper, lateral walls of the fourth ventricular cavity, and the inferior cerebellar peduncle (ICP) forms the walls elsewhere. At the lateral recesses of the ventricle, near the foramina of Luschka, the vestibular and cochlear nerves enter. In the ventricular floor, there are longitudinal fissures or sulci separating ridges and protuberances. The medial longitudinal fissure lies in the midline and separates the two sides. The paired grooves of the sulcus limitans, separating basal plate (motor) structures from alar plate (sensory) structures, lie laterally.
The striae medullares of the fourth ventricle (stria medullares medullares) is a band of myelinated fibers running across the ventricular floor. The fibers arise from the external arcuate nucleus, which lies anterior to the medullary pyramids, and are bound for the ICP. Paired midline humps in the ventricular floor rostral to the stria medullares are the facial colliculi, beneath which are the nuclei of CN VI and the internal genu of CN VII. Along the same meridians caudal to the striae are the hypoglossal trigones, beneath which lie the nuclei of CN XII. Lateral to the hypoglossal trigones are the vagal trigones (ala cinerea), beneath which are the dorsal motor nuclei of the vagus nerves. The area postrema (chemoreceptor trigger zone) is a narrow strip along the caudal aspect of the vagal trigone. Far laterally, near the entry zones of CN VIII, are the vestibular areas. At the caudal tip of the fourth ventricle is the obex, the point at which the fourth ventricle communicates with the central canal of the spinal cord. The shape of the rhomboid fossa at the caudal end of the ventricle resembles a writing pen; it is referred to as the calamus scriptorius. On the dorsal surface caudal to the ventricle are the gracile tubercles in the midline and the cuneate tubercles just laterally; these merge into the gracile and cuneate fasciculi inferiorly. Lateral to the gracile and cuneate tubercles are the ICPs.
Brainstem Organization
The brainstem, throughout its length, is composed of three parts: tectum (roof), tegmentum (midportion), and base (Figure 11.5). In the midbrain, the tectum consists of the quadrigeminal plate. In the pons and medulla, the tectum devolves into nonfunctional tissue forming the roof plate of the fourth ventricle, the anterior (superior) medullary velum in the pons, and the posterior (inferior) medullary velum in the medulla. The contents of the tegmentum are variable from level to level, and include the CN motor and sensory nuclei. Running throughout the length of the tegmentum is the RF. The reticular activating system is part of this loose network and is responsible for controlling arousal. Coursing through the tegmentum are the long ascending and descending tracts (e.g., medial lemniscus (ML), spinothalamic tract, rubrospinal tract, and others). The base consists of descending corticospinal and corticobulbar fibers in different configurations.
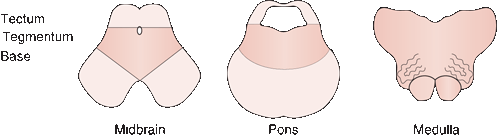
FIGURE 11.5 Three levels of the brainstem showing what constitutes the tectum, tegmentum, and base at each level. (Reprinted from Campbell WW, Pridgeon RM. Practical Primer of Clinical Neurology. Philadelphia: Lippincott Williams & Wilkins, 2002, with permission.)
Reticular Formation
The core of the brainstem is the RF a loose network of cells and fibers that has extensive interconnections with other brainstem structures as well as complex, polysynaptic projections rostrally and caudally. The RF terminates as the intralaminar nuclei of the thalamus. There are three cell populations in the RF: the raphe nuclei, the medial reticular nucleus, and the lateral reticular nucleus. The raphe nuclei are a detached series of individual nuclear groups that lie in the midline (Gr. raphe “seam”) from the rostral midbrain to the caudal medulla. All the raphe nuclei send serotonergic projections widely throughout the nervous system. As a generalization, the midbrain raphe nuclei project to the hemispheres, those in the pons to the brainstem and cerebellum, and those in the medulla to the spinal cord.
The lateral reticular nucleus contains small neurons and is primarily afferent; it receives collateral projections from ascending and descending long tracts. These parvocellular neurons are essentially a continuation of the system of interneurons in the spinal cord. The lateral reticular nucleus projects primarily to the medial reticular nucleus. The cells of the medial reticular nucleus are larger, and these magnocellular neurons send projections up and down the neuraxis. An expansion of the nucleus in the upper medulla forms the medullary gigantocellular nucleus, and in the pons the pontine gigantocellular nucleus. The medial reticular nucleus gives rise to two major descending tracts. The medial reticulospinal (bulbospinal) tract arises from the medullary nucleus and the lateral reticulospinal (pontospinal) tract from the pontine nucleus.
Brainstem Nuclei
The major brainstem nuclei are depicted in Figure 11.6. Some exist as focal collections, others as cell columns that range longitudinally over an extensive span. The location, composition, and function of these nuclei are summarized in Tables 11.1 and 11.2.
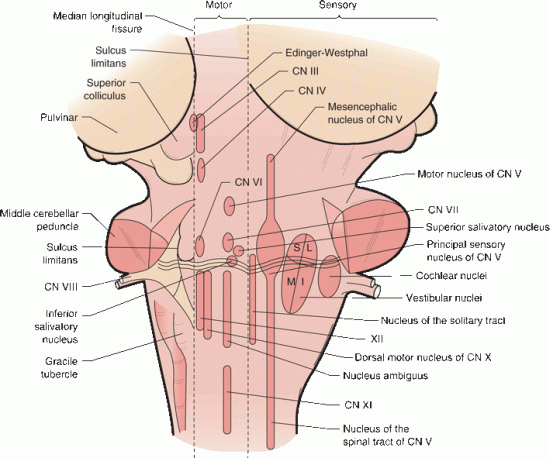
FIGURE 11.6 Gross representational brainstem view on the left demonstrates the relationships of the external structures. On the right, the expanded diagrammatic view shows the location of the various cell columns that lie interiorly, motor nuclei medially, and sensory nuclei laterally, separated by the sulcus limitans.




