CHAPTER 8
Cranial Nerves and Pathways
The 12 pairs of cranial nerves are referred to by either name or Roman numeral (Fig 8–1 and Table 8–1). Note that the olfactory peduncle (see Chapter 19) and the optic nerve (see Chapter 15) are not true nerves but rather fiber tracts of the brain, whereas nerve XI (the spinal accessory nerve) is derived, in part, from the upper cervical segments of the spinal cord. The remaining nine pairs relate to the brain stem.
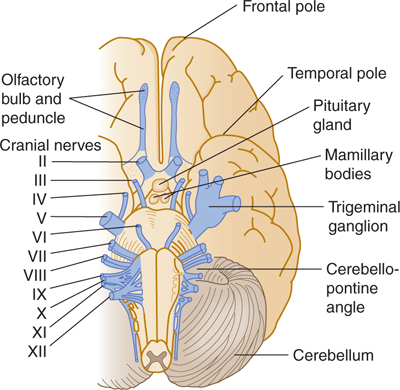
FIGURE 8–1 Ventral view of the brain stem with cranial nerves.
Table 8–1 Overview of Cranial Nerves.
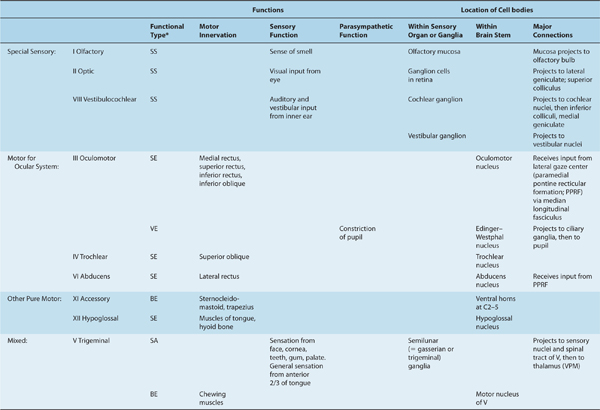

ORIGIN OF CRANIAL NERVE FIBERS
Cranial nerve fibers with motor (efferent) functions arise from collections of cells (motor nuclei) that lie deep within the brain stem; they are homologous to the anterior horn cells of the spinal cord. Cranial nerve fibers with sensory (afferent) functions have their cells of origin (first-order nuclei) outside the brain stem, usually in ganglia that are homologous to the dorsal root ganglia of the spinal nerves. Second-order sensory nuclei lie within the brain stem (see Chapter 7 and Fig 7–6).
Table 8–1 presents an overview of the cranial nerves. This table does not list the cranial nerves numerically; rather, it groups them functionally:
 Nerves I, II, and VIII are devoted to special sensory input.
Nerves I, II, and VIII are devoted to special sensory input.
 Nerves III, IV, and VI control eye movements and pupillary constriction.
Nerves III, IV, and VI control eye movements and pupillary constriction.
 Nerves XI and XII are pure motor (XI: sternocleidomastoid and trapezius; XII: muscles of tongue).
Nerves XI and XII are pure motor (XI: sternocleidomastoid and trapezius; XII: muscles of tongue).
 Nerves V, VII, IX, and X are mixed.
Nerves V, VII, IX, and X are mixed.
 Note that nerves III, VII, IX, and X carry parasympathetic fibers.
Note that nerves III, VII, IX, and X carry parasympathetic fibers.
FUNCTIONAL COMPONENTS OF THE CRANIAL NERVES
A cranial nerve can have one or more functions (as shown in Table 8–1). The functional components are conveyed from or to the brain stem by six types of nerve fibers:
(1) Somatic efferent fibers, also called general somatic efferent fibers, innervate striated muscles that are derived from somites and are involved in eye (nerves III, IV, and VI) and tongue (nerve XII) movements.
(2) Branchial efferent fibers, also known as special visceral efferent fibers, are special somatic efferent components. They innervate muscles that are derived from the branchial (gill) arches and are involved in chewing (nerve V), making facial expressions (nerve VII), swallowing (nerves IX and X), producing vocal sounds (nerve X), and turning the head (nerve XI).
(3) Visceral efferent fibers are also called general visceral efferent fibers (preganglionic parasympathetic components of the cranial division); they travel within nerves III (smooth muscles of the inner eye), VII (salivatory and lacrimal glands), IX (the parotid gland), and X (the muscles of the heart, lung, and bowel that are involved in movement and secretion; see Chapter 20).
(4) Visceral afferent fibers, also called general visceral afferent fibers, convey sensation from the alimentary tract, heart, vessels, and lungs by way of nerves IX and X. A specialized visceral afferent component is involved with the sense of taste; fibers carrying gustatory impulses are present in cranial nerves VII, IX, and X.
(5) Somatic afferent fibers, often called general somatic afferent fibers, convey sensation from the skin and the mucous membranes of the head. They are found mainly in the trigeminal nerve (V). A small number of afferent fibers travel with the facial (VII), glossopharyngeal (IX), and vagus (X) nerves; these fibers terminate on trigeminal nuclei in the brain stem.
(6) Special sensory fibers are found in nerves I (involved in smell), II (vision), and VIII (hearing and equilibrium).
Differences Between Cranial and Spinal Nerves
Unlike the spinal nerves, cranial nerves are not spaced at regular intervals. They differ in other aspects as well: The spinal nerves, for example, contain neither branchial efferent nor special sensory components. Some cranial nerves contain motor components only (most motor nerves have at least a few proprioceptive fibers), and some contain large visceral components. Other cranial nerves are completely or mostly sensory, and still others are mixed, with both types of components. The motor and sensory axons of mixed cranial nerves enter and exit at the same point on the brain stem. This point is ventral or ventrolateral except for nerve IV, which exits from the dorsal surface (see Fig 8–1).
The optic nerve is unique in that it connects the retina (which some nerves scientists consider a specialized outpost of the brain) with the brain. The optic nerve is essentially a white matter tract that connects the retina to the brain. Axons within the optic nerve are myelinated by oligodendrocytes, in contrast to axons within peripheral nerves that are myelinated by Schwann cells.
Ganglia Related to Cranial Nerves
Two types of ganglia are related to cranial nerves. The first type contains cell bodies of afferent (somatic or visceral) axons within the cranial nerves. (These ganglia are somewhat analogous to the dorsal root ganglia that contain the cell bodies of sensory axons within peripheral nerves.) The second type contains the synaptic terminals of visceral efferent axons, together with postsynaptic (parasympathetic) neurons that project peripherally (Table 8–2).
Table 8–2 Ganglia Related to Cranial Nerves.
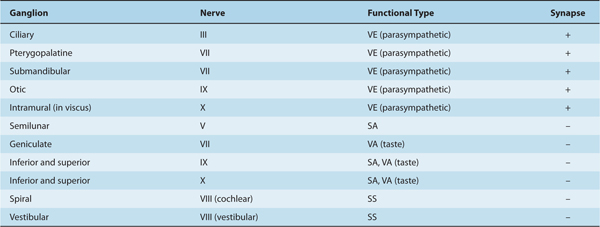
Sensory ganglia of the cranial nerves include the semilunar (gasserian) ganglion (nerve V), geniculate ganglion (nerve VII), cochlear and vestibular ganglia (nerve VIII), inferior and superior glossopharyngeal ganglia (nerve IX), superior vagal ganglion (nerve X), and inferior vagal (nodose) ganglion (nerve X).
The ganglia of the cranial parasympathetic division of the autonomic nervous system are the ciliary ganglion (nerve III), the pterygopalatine and submandibular ganglia (VIII), otic ganglion (IX), and intramural ganglion (X). The first four of these ganglia have a close association with branches of V; the trigeminal branches may course through the autonomic ganglia.
ANATOMIC RELATIONSHIPS OF THE CRANIAL NERVES
Cranial Nerve I: Olfactory Nerve
The true olfactory nerves are short connections that project from the olfactory mucosa within the nose and the olfactory bulb within the cranial cavity (Fig 8–2; see also Chapter 19). There are 9 to 15 of these nerves on each side of the brain. The olfactory bulb lies just above the cribriform plate and below the frontal lobe (nestled within the olfactory sulcus). Axons from the olfactory bulb run within the olfactory stalk, synapse in the anterior olfactory nucleus, and terminate in the primary olfactory cortex (pyriform cortex) as well as the entorhinal cortex and amygdala.
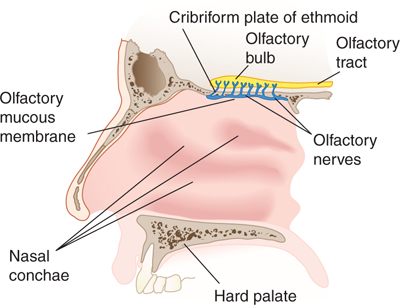
FIGURE 8–2 Lateral view of the olfactory bulb, tract, mucous membrane, and nerves.
Cranial Nerve II: Optic Nerve
The optic nerve contains myelinated axons that arise from the ganglion cells in the retina. As noted above, axons within the optic nerve are myelinated by oligodendrocytes. The optic nerve passes through the optic papilla to the orbit, where it is contained within meningeal sheaths. The nerve changes its name to optic tract when the fibers have passed through the optic chiasm (Fig 8–3). Optic tract axons project to the superior colliculus and to the lateral geniculate nucleus within the thalamus, which relays visual information to the cortex (see Chapter 15).
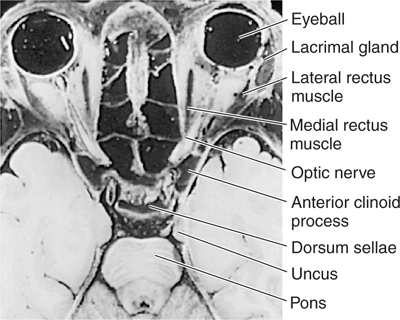
FIGURE 8–3 Horizontal section through the head at the level of the orbits.
Cranial Nerve III: Oculomotor Nerve
Cranial nerves III, IV, and VI work together to control eye movements and are therefore discussed together. In addition, cranial nerve III controls pupillary constriction.
The oculomotor nerve (cranial nerve III) contains axons that arise in the oculomotor nucleus (which innervates all of the oculomotor muscles except the superior oblique and lateral rectus) and the nearby Edinger-Westphal nucleus (which sends preganglionic parasympathetic axons to the ciliary ganglion). The oculomotor nerve leaves the brain on the medial side of the cerebral peduncle, behind the posterior cerebral artery and in front of the superior cerebellar artery. It then passes anteriorly, parallel to the internal carotid artery in the lateral wall of the cavernous sinus, leaving the cranial cavity by way of the superior orbital fissure.
The somatic efferent portion of the nerve innervates the levator palpebrae superioris muscle; the superior, medial, and inferior rectus muscles; and the inferior oblique muscle (Fig 8–4). The visceral efferent portion innervates two smooth intraocular muscles: the ciliary and the constrictor pupillae.
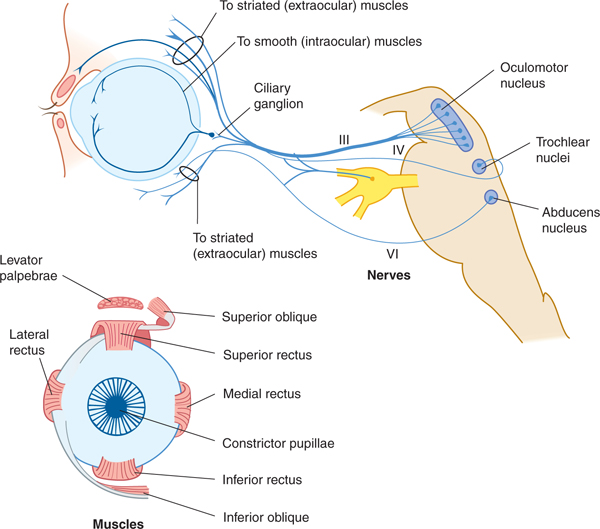
FIGURE 8–4 The oculomotor, trochlear, and abducens nerves; ocular muscles.
CLINICAL CORRELATIONS
Cranial Nerve IV: Trochlear Nerve
The trochlear nerve is the only crossed cranial nerve. It originates from the trochlear nucleus, which is a group of specialized motor neurons located just caudal to (and actually constituting a subnucleus of) the oculomotor nucleus within the lower midbrain. Trochlear nerve axons arise from these neurons, cross within the midbrain, and then emerge contralaterally on the dorsal surface of the brain stem. The trochlear nerve then curves ventrally between the posterior cerebral and superior cerebellar arteries (lateral to the oculomotor nerve). It continues anteriorly in the lateral wall of the cavernous sinus and enters the orbit via the superior orbital fissure. It innervates the superior oblique muscle (see Fig 8–4).
Note: Because nerves III, IV, and VI are generally grouped together for discussion, nerve V is discussed after nerve VI.
Cranial Nerve VI: Abducens Nerve
A. Anatomy
The abducens nerve arises from neurons of the abducens nucleus located within the dorsomedial tegmentum within the caudal pons. These axons project through the body of the pons and leave it as the abducens nerve. This nerve emerges from the pontomedullary fissure, passes through the cavernous sinus close to the internal carotid, and exits from the cranial cavity via the superior orbital fissure. Its long intracranial course makes it vulnerable to pathologic processes in the posterior and middle cranial fossae. The nerve innervates the lateral rectus muscle (see Fig 8–4).
A few sensory (proprioceptive) fibers from the muscles of the eye are present in nerves III, IV, and VI and in some other nerves that innervate striated muscles. The central
B. Action of the External Eye Muscles
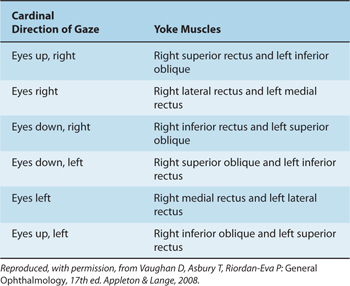
The actions of eye muscles operating singly and in tandem are shown in Tables 8–3 and 8–4 (Fig 8–5). The levator palpebrae superioris muscle has no action on the eyeball but lifts the upper eyelid when contracted. Closing the eyelids is performed by contraction of the orbicular muscle of the eye; this muscle is innervated by nerve VII.
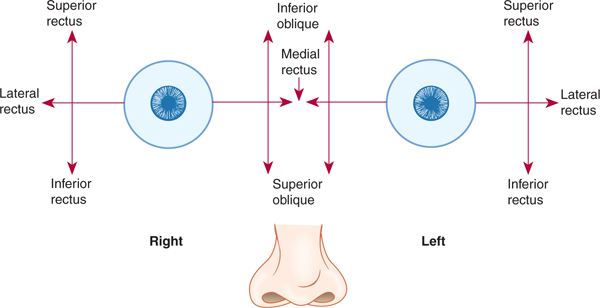
FIGURE 8–5 Diagram of eye muscle action.
Table 8–3 Functions of the Ocular Muscles.
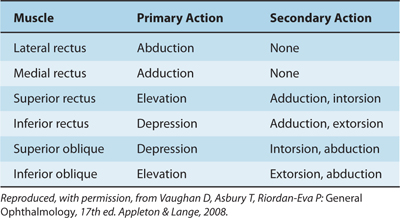
Table 8–4 Yoke Muscle Combinations.
C. Control of Ocular Muscle Movements
The oculomotor system activates the various extraocular muscles in a highly coordinated manner (Fig 8–6). When the eyes scan the environment, they do so in short, rapid movements called saccades. When a target moves, a different form of ocular movement—smooth pursuit—is used to keep the image in sharp focus. When the head or body moves unexpectedly (eg, when one is jolted), reflex movements of the head and eye muscles compensate and maintain fixation on the visual target. This compensatory function is achieved by the vestibuloocular reflex (see Chapter 17).
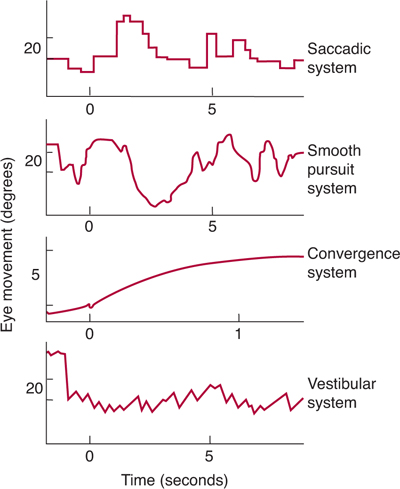
FIGURE 8–6 Types of eye movement control. (Modified and reproduced, with permission, from Robinson DA: Eye movement control in primates. Science 1968;161:1219. Copyright ©1968 by the American Association for the Advancement of Science.)
The six individual muscles that move one eye normally act together with the muscles of the other eye in controlled movement. Both eyes move in the same direction to follow an object in space, but they move by simultaneously contracting and relaxing different muscles; this is called a conjugate gaze movement. Fixating on a single point is called vergence, which requires a different set of muscles, including the intraocular muscles. Each of the extraocular muscles is brought into play in conjugate gaze movements or vergence.
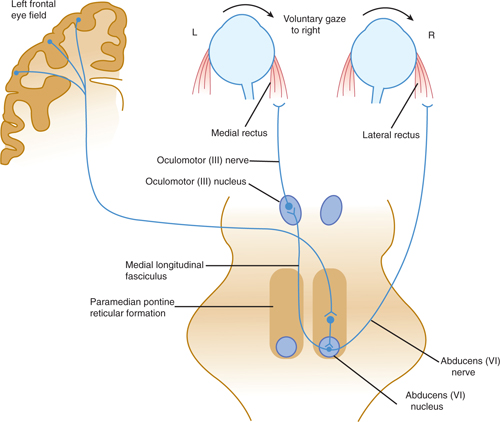
1. Gaze and vergence centers—Conjugate gaze and vergence are controlled from three areas in the brain stem. There are two lateral gaze centers in the paramedian pontine reticular formation near the left and right abducens nuclei and a vergence center in the pretectum just above the superior colliculi. Each of these three areas can be activated during head movement by the vestibular system via the medial longitudinal fasciculus (see Chapter 17). Activation of the lateral gaze center on the right produces conjugate gaze to the right and vice versa. Regions in the contralateral frontal lobe (the eye field area) influence voluntary eye movements via polysynaptic connections to the lateral gaze centers, whereas regions in the occipital lobe influence visual pursuit and also have connections with the vergence center (Fig 8–7).
Activity in each of the lateral gaze centers (located in the paramedian pontine reticular formation on each side, adjacent to the abducens nuclei) controls eye movements to the ipsilateral side. Thus, the lateral gaze center on the right is connected, via excitatory projections, to the right abducens nucleus that activates the lateral rectus muscle responsible for abduction of the right eye. The right-sided lateral gaze center also sends projections, via the medial longitudinal fasciculus, to the contralateral (left-sided) oculomotor nucleus, where they form excitatory synapses on oculomotor neurons innervating the medial rectus muscle. (This muscle is responsible for movement of the left eye across the midline to the right.) As a result of this arrangement, activation of the right-sided lateral gaze center results in movement of both eyes to the right (see Fig 8–7).
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


