CHAPTER 4 DISORDERS OF MEMORY
DEFINITIONS AND TERMINOLOGY
Declarative and Nondeclarative Memory
In general, when patients speak of memory complaints, they are referring to the recollection of information for which there is conscious awareness. In neuropsychological terminology, this is called declarative, or explicit, memory. The corollary is nondeclarative, or implicit, memory, which refers to phenomena such as motor skill learning, priming and classical conditioning (Table 4-1). For instance, with appropriate training, a tennis player can be seen to “learn,” as evidenced by improved performance; however, in the course of a rally, the player is not consciously recollecting the motor sequence required to execute each shot. Priming refers to a situation in which prior exposure leads to altered performance and can be shown experimentally in language tasks such as naming and lexical decision making. For example, imagine being presented with words on a computer screen and being asked to read them as soon as they appear; the latency between exposure and response (reaction time) is shorter if a related, as opposed to a nonrelated, item is presented immediately before the test item (e.g., a subject will respond faster to the word tiger if it is preceded by lion rather than house). Highlighting the dissociation between declarative and nondeclarative memory is the fact that patients with Alzheimer’s disease have marked declarative deficits and yet may respond even faster with priming (hyperpriming) than control subjects. Although nondeclarative memory is crucial for many functions, it is not what patients usually mean when they report memory symptoms and is not discussed further.
TABLE 4-1 Classification of Memory Types
Episodic and Semantic Memory
Declarative memory is also considered in terms of encoding, storage, and retrieval. In other words, a neurophysiological change takes place during learning (encoding); this change must leave some enduring trace (storage), and there must be a mechanism by which this trace can be reactivated, at will, to lead to the subjective experience of remembering (retrieval). Often, these processes cannot be disentangled in the clinic; for instance, a patient with no recall of new information on formal neuropsychological testing may have a deficit at any or all of these stages. Nevertheless, neuropsychological tests can, to some extent, tease these stages apart by varying task demands. For example, a patient who requires an excessive number of learning trials to reach a criterion and yet retains a lot of this information after a delay can be considered as having an encoding problem. This profile is often indicative of an attention disorder rather than a true amnesic syndrome. Storage problems may be suggested by an accelerated decay in recall performance between two time points (e.g., immediately and 30 minutes after encoding). However, it is important to realize that a degree of decay is seen under normal circumstances; therefore, interpretation requires comparison with demographically matched norms. Retrieval deficits can be investigated by comparing free recall (“What did I ask you to remember?”) with recognition memory (“Which of these did I show you earlier?”). Recognition is typically tested by either forced-choice questions—in which target answers and foils (incorrect alternatives) are presented simultaneously in pairs and the patient has to choose those that he or she has seen before—or by asking for yes/no responses as targets and foils are presented in a random sequence. Because the patient is presented the previously studied material, retrieval demands are minimized, and hence a retrieval deficit is suggested where there is disproportionate impairment of free recall in comparison with recognition memory. Recognition is, however, an easier task than is free recall, and so this dissociation also needs to be assessed by comparison with control norms. It is likely that the neural systems responsible for these processes partially overlap, in that some brain areas such as the hippocampus may be necessary for all aspects, whereas other areas may be differentially engaged at a specific stage. For example, functional activation studies of changes in cerebral blood flow suggest greater engagement of the left and right prefrontal cortices during encoding and retrieval, respectively.1
Amnesia
Amnesia is the generic term for loss of memory. An inability to establish new memories after the pathological event is called anterograde amnesia, whereas the inability to recall memories that had been established before the pathological event is called retrograde amnesia (Fig. 4-1). The fact that the distinction between anterograde and retrograde is referenced to the pathological insult is important. Students often think erroneously that retrograde amnesia refers to memory loss for past events, whereby “past” is loosely defined as any time before the clinical consultation. This leads to the absurd situation of testing for retrograde deficits by asking the patient what they did “yesterday,” which makes the distinction from anterograde deficits meaningless, as it becomes referenced to the ever-changing present. Patients with dense amnesia, especially in the acute phase, may confabulate: that is, fill in memory gaps with false statements.
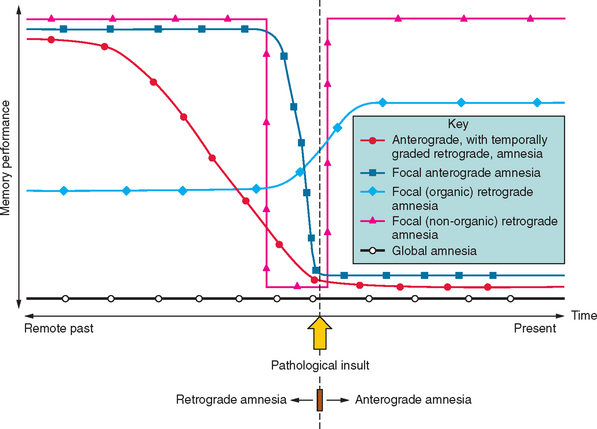
The situation becomes more ambiguous with regard to factual semantic knowledge (public events, famous people), including personal semantic facts (e.g., names of schools attended, name of employers). Variable degrees of impairment in factual semantic knowledge are usual in amnesia, and this information can be objectively dated in time (in contrast to an individual’s episodic recollections, which are more difficult to date or verify). As a consequence, evidence for temporal gradients in memory impairment is typically compiled from these datable facts. For instance, a patient with a new-onset pathological process who knows that the Berlin Wall was pulled down, that apartheid ended, and that Iraq invaded Kuwait but does not remember a tsunami killing hundreds of thousands, the World Trade Center being destroyed, or Princess Diana dying has evidence for a retrograde amnesia extending back to the early to mid-1990s. It should be noted that the distinction between episodic and semantic memory—the degree of “semanticization”—becomes blurred at this point, inasmuch as some facts may be recalled from the specific context in which they were encountered—as so-called “flashbulb” memories. In addition, the frequency of exposure to particular facts varies between individuals. For example, the average American has most likely had many more encounters with stories about the World Trade Center than with stories about the Berlin Wall, and this will bias successful recollection towards the more recent, semantically reinforced event.
Working Memory
An important concept to distinguish from both episodic and semantic memory is working memory, frequently referred to by experimental psychologists as short-term memory. This refers to information that, at any given moment, is held “on-line” by the brain—that is, in consciousness—and that is not recollected after a distraction period (unless incidental episodic encoding of the material has also taken place). This faculty is exemplified by the ability to keep a telephone number in the mind in the interval between looking it up the telephone directory and dialing the number. Models of working memory posit auditory-verbal and visuospatial components that are coordinated and manipulated by a central executive system.2 A highly distributed network of cortical and subcortical areas supports working memory, although the dorsolateral prefrontal cortex appears to be particularly important for the executive component. It is therefore unsurprising that working memory is most vulnerable to diffuse insults such as those found in metabolic encephalopathies and closed head injury. It can be dissociated from the long-term declarative memory systems described previously; patients with even the densest amnesia can still have intact working memory.
NEUROBIOLOGY OF LEARNING AND MEMORY
Cellular Mechanisms
These observations suggest that the transmission properties of existing synapses can be modified by usage, but it is notable that the “hard wiring” of the nervous system is also dynamic. The dendritic spines, the sites of synaptic connections in the neuronal dendritic arbor, also display plasticity over time, which suggests that the neural networks themselves may be adaptive.3 Technical developments, enabling dendritic spines to be labeled and studied in real time, have shown that changes in structure, including the generation of new dendritic spine protrusions, can be triggered by LTP-inducing activity. Conversely, some LTD-inducing stimuli have been shown to cause loss of dendritic spines. It must be acknowledged, however, that although these morphological and physiological findings suggest mechanisms by which learning and memory may be supported, the precise relevance of these specific processes to memory in vivo is far from established in the mature brain.
For further reading on these topics and other current issues in the neuroscience of memory, the excellent essays by Dudai (2002) are recommended.
The Topography of Memory
Between synaptic plasticity and the topographical anatomy of regions implicated in amnesia, the precise nature of the memory trace remains difficult to delineate. As already discussed, the information that constitutes episodic memory comprises multiple sensory modalities. The prevailing hypothesis is that structures such as the hippocampus do not “store” this information, but rather act to bind the disparate components that constitute a memory trace. To illustrate this model, consider an episodic memory of a specific conversation comprising, for simplicity, an auditory component (the dialogue) and a visual component (the scene). At the time of encoding, visual and auditory inputs contemporaneously activate a neural network in the isocortex, including auditory and visual association cortex, as well as the mesial temporal lobe. The temporospatial firing pattern that represents the memory trace can then be subsequently reactivated via the hippocampus. The traditional view (Fig. 4-2) is that with time, this neural representation gradually becomes independent of the hippocampus: so-called consolidation. This is the explanation for the Ribot effect, a temporal gradient in retrograde amnesia in which amnesia is most dense for the time immediately before the pathological insult and is progressively less dense for earlier periods. This view is almost certainly an oversimplification; many amnesic patients do not exhibit a Ribot effect but, rather, a flat profile of memory loss extending back over their whole life. Furthermore, when it is found, the temporal gradient often extends back over several decades, which suggests that consolidation is an implausible entity in evolutionary terms, inasmuch as it implies that in old age, the brain is still consolidating memories from early adult life.4
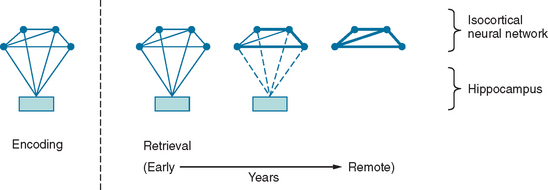
Another hotly debated topic is whether the distinction between semantic and episodic memory is meaningful in biological terms. Evidence for two separable memory systems comes from the observation that individuals with episodic memory amnesia have preservation of general semantic knowledge. Furthermore, neuropsychological studies have shown that patients with hippocampal amnesia, including those with damage sustained in infancy,5 can acquire new semantic knowledge in spite of the amnesia. This suggests that semantic memory, including its acquisition, can be supported independently of the hippocampus. The opposing view is that the acquisition of both semantic and episodic memory is dependent on a unitary system. Evidence for this comes from the observation that although acquisition of semantic knowledge may occur in patients with hippocampal amnesia, it is not completely normal.6 Whereas each episodic memory is unique in time and space, semantic facts are experienced in multiple contexts. This means that there is an enormous encoding advantage for the latter, which may explain the dissociation.
In addition to the interaction of the hippocampus and isocortical sensory association areas in sustaining declarative memory, the amygdala and prefrontal cortex merit mention. The amygdala is involved in emotional processes, such as recognition of fear in other’s faces, but with regard to memory, it is thought to reinforce encoding of emotionally salient events. Clearly, people do not recollect their entire life experience as a bland, continuous narrative. The fact that some experiences are forgotten but others are remembered is related in part to their differing emotional significance, and it is thought that encoding of emotionally significant events is facilitated by an interaction of the amygdala with other mesial temporal structures.7 The prefrontal cortex is thought to have a “meta” role in mnemonic processing, being involved in focusing attention for encoding, forming retrieval strategies, and monitoring output.8 For instance, when a person is asked to produce a list of animals, performance is enhanced if the list is clustered into semantic categories (“category clustering”) such as zoo animals, farm animals, and domestic animals. The prefrontal cortex is specifically engaged in this type of processing, but not in storing the memories per se. Consequently, frontal lesions can cause deficits in free recall with relative preservation of recognition, because the latter does not require an active retrieval strategy.
ASSESSMENT OF MEMORY
Clinical Assessment
Wherever possible, a history should be obtained from a close relative of the patient to cross-reference to the patient’s account. After the clinician explains the procedure to the patient and obtains his or her consent, the informant should be seen alone, to allow frank discussion without fear of embarrassment. A common mistake in assessing patients with memory symptoms is failure to examine memory adequately. There is often insufficient time in a general clinic to test delayed recall over a long interval; however, new learning can be informally assessed by giving the patient some material to encode and then testing recall and recognition after a 5-minute distraction period filled with other elements of the examination. A seven-item name and address recall can be assessed without any special equipment; this test yields indices of encoding, recall, and recognition (Fig. 4-3). Asking the patient to copy abstract designs, followed by testing of spontaneous recall and recognition, can help assess nonverbal memory.

Neuropsychological Assessment
The neuropsychologist also documents the patient’s history, including the demographic information necessary for test interpretation (age, education, handedness). Memory performance also needs to be interpreted in the context of general cognitive abilities; thus, the assessment is never restricted to memory alone. A vast array of tests are available that probe various aspects of memory: encoding, retrieval, and recognition; verbal and nonverbal memory; remote memory; and so on. Some of the memory tests in common usage are listed in Table 4-2. Reliable interpretation of neuropsychological performance requires that the tests be administered under standardized conditions by an appropriately trained examiner. Ad hoc incorporation of neuropsychological tests into the bedside examination is therefore best avoided.
TABLE 4-2 Selected Neuropsychological Tests of Memory
| Test | Description | Reference or Source |
|---|---|---|
| New Learning | ||
| Rey Auditory Verbal Learning Test (RAVLT) | Word list encoding, free recall, and recognition | Rey A: L’Examen Clinique en Psychologie. Paris: Presses Universitaires de France, 1964 |
| California Verbal Learning Test (CVLT) | Word list semantic clustering, encoding, free recall and recognition | Delis DC, Kaplan E, Kramer J, et al: California Verbal Learning Test (CVLT-II) Second Edition—Adult Version. San Antonio, TX: The Psychological Corporation, 2000 |
| Story Recall (Logical Memory) | Story free recall ± recognition | Wechsler D: Wechsler Memory Scale—Revised. San Antonio, TX: The Psychological Corporation, 1987. |
| Recognition Memory Test (RMT) | Face and word recognition | Warrington, EK: Recognition Memory Test. Windsor, UK: NFER-Nelson, 1984 |
| Doors and People Test | Verbal and nonverbal recall and recognition | Baddeley A, Emslie H, Nimmo-Smith I: Doors and people. Oxford, UK: Thames Valley Test Company, 1994 |
| Rey-Osterrieth Complex Figure* | Nonverbal recall ± recognition | Osterrieth PA: Le Test de Copie d’Une Figure Complex [Complex Figure Copy Test]. Arch Psychol 1944; 30:206-356. |
| Paired Associates Learning (PAL) | Object/spatial location recall | Cambridge Neuropsychological Test Automated Battery. Cambridge, UK: Cambridge Cognition, Ltd. |
| Rivermead Behavioural Memory Test (RBMT) | Various tests including some “real-life” memory tasks | Wilson BA, Cockburn J, Baddeley A: Rivermead Behavioural Memory Test (RBMT-II). Oxford, UK: Thames Valley Test Company, 2003 |
| Remote Memory | ||
| Autobiographical Memory Interview (AMI) | Personal semantics and episodes from childhood, early adulthood, and recent past | Kopelman M, Wilson B, Baddeley A: The Autobiographical Memory Interview (AMI). Oxford, UK: Thames Valley Test Company, 1990 |
| Galton-Crovitz cue-word test | Specific autobiographical episodes generated to word prompts | Crovitz HF, Schiffman H: Frequency of episodic memories as a function of their age. Bull Psychonom Soc 1974; 4:517-518 |
| General Semantic Memory | ||
| Category fluency | Generation of exemplars to a target category | |
| Boston Naming Test | Picture naming | Kaplan E, Goodglass H, Weintraub S: Boston Naming Test. Philadelphia: Lea & Febiger (Also published by Psychcorp and LWW.), 1983 |
| Graded Naming Test | Picture naming | McKenna P, Warrington EK: The Graded Naming Test. Windsor, UK: NFER-Nelson, 1993 |
| Pyramids and Palm Trees Test | Word and picture forced-choice associative knowledge | Howard D, Patterson K: Pyramids and Palm Trees Test. Oxford, UK: Thames Valley Test Company, 1992 |
| Factual Semantic Memory | ||
| Variations on tests of famous people, news events, etc.† | ||
† These tests are typically used to assess the temporal extent of amnesia; however, because knowledge of such events varies greatly between communities, these tests are typically devised by individual neuropsychology laboratories.
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


