CHAPTER 5 HIGHER VISUOPERCEPTUAL DISORDERS AND DISORDERS OF SPATIAL COGNITION
At least one third of the primate brain is devoted to visual perception and visual processing.1 A vast amount has been learned about the visual system in the past 100 years or so, from both laboratory studies and studies of clinical and healthy populations. This research has identified multiple areas in the cerebral cortex (not just the occipital lobe) that receive visual input (Figs. 5-1 through 5-3). There is both parallel and distributed processing of visual stimuli. One of the most famous depictions are the diagrams of Van Essen and colleagues,2 which map the macaque cerebral cortex (Fig. 5-1).
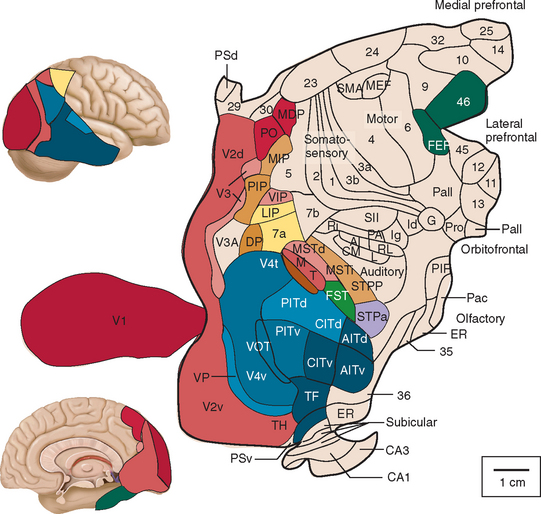
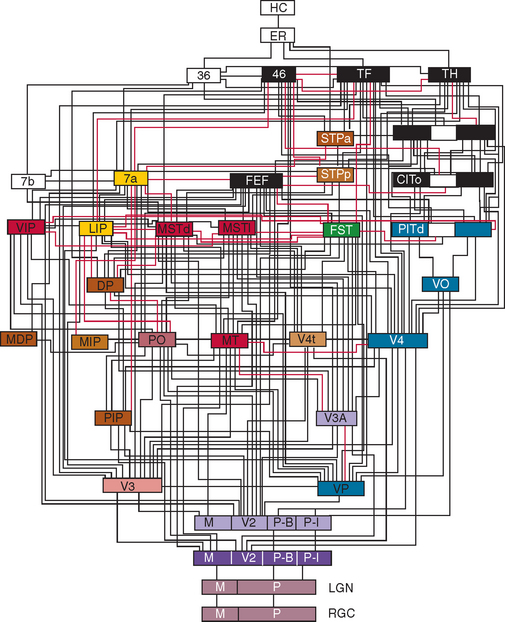
Figure 5-2 Hierarchy of visual areas in the macaque. This diagram shows over 30 visual cortical areas, shaded similarly to Figure 5-1, two subcortical visual stages—the retinal ganglion cell layer (RGC) and the lateral geniculate nucleus (LGN), plus several non-visual areas such as area 7b of the somatosensory cortex, area 36 and the hippocampal complex (HC). These areas are connected by extensive linkages, most of which have been demonstrated to be reciprocal pathways. The patterns of connections help to illustrate two fundamental principles of organization: hierarchical connections and parallel processing.
(From Felleman DJ, Van Essen DC: Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex 1991; 1:1-47.)

Many disorders of higher visuoperceptual function have been described, some of them in syndromic fashion, and include Balint’s syndrome, cerebral akinetopsia (rare), cerebral dyschromatopsia (somewhat more common), disorders of face recognition (prosopagnosia), environmental disorientation (which overlaps with spatial disorientation: see also Chapter 6), and visual agnosias. Even one of the rarer language disorders may be thought of as a primary visual disturbance: alexia without agraphia (see Chapter 3). Higher-order visual complaints are often one of the first indications of a number of other neurological illnesses such as prion diseases.
EPIDEMIOLOGY
Many of the diseases that underlie these abnormalities are age related—in particular, the dementias and cerebrovascular disease—so these entities are more frequent in older persons and are increasingly commonly encountered as the population ages. Within the neurodegenerative disorders, using Alzheimer’s disease as the prototype, estimates for the prevalence of higher-order visual disturbances are as high as 57%, mainly because of a high prevalence of visual agnosia. The severity of the dementia is strongly correlated with the complexity of the presenting visual disturbance.3
The effects of these higher visuoperceptual disorders are amplified by other disorders of perception that may be present in older people, including visual loss because of anterior visual pathway disease (e.g., cataract, macular degeneration) and impairments of hearing and mobility. Studies have suggested that of people between the ages of 75 and 79, at least 12% have some visual impairment, rising to 23% in the 85- to 89-year-old age range.4 By 90 years of age, at least one in three people will have some visual deficit.4,5
THE TWO-SYSTEMS APPROACH TO HIGHER VISUOSPATIAL FUNCTIONING
Higher visual processing attempts to achieve two primary objectives: the identification of visual stimuli and their localization in space. These two goals of visual analysis are often abbreviated as “what” and “where” (Fig. 5-4). Research has demonstrated that these two goals are achieved relatively independently through two anatomically separated systems known as the ventral (what) and dorsal (where) visual processing pathways.6–8 The fundamental concept is that there is a dorsal stream of information concerned with an object’s location and a ventral stream concerned with its identity. Because this distinction is so widespread in the literature, it must be considered, but even the proponents of this dualistic model freely admit that it is an oversimplification of the true situation, given that strong interactions occur between the two networks.

ANATOMY AND PHYSIOLOGY
In general, higher visuoperceptual and visuospatial disorders reflect damage to one or several multiple brain regions including (but not restricted to) the occipital lobes, temporal lobes, parietal lobes, and underlying white matter. In many of the described syndromes, the right hemisphere appears to be more affected than the left, with many investigators proposing that much of the bilateral activation on higher visual tasks results from the transcallosal influence of the right hemisphere.9,10 The precise nature of the damage for each particular disorder is discussed in the following sections.
Disorders of the Temporal Vision–Related Cortex (Ventral Processing Stream)
Damage to the cortical regions of the ventral processing stream results in a variety of disturbances of visual form and color perception (Table 5-1). These can include dyschromatopsia and alexia without agraphia. Most commonly, however, damage to these regions results in a form of visual agnosia, a clinical syndrome characterized by an inability to recognize a visually presented object despite the presence of adequate cognition, visual acuity, attention, and language skills.11,12
TABLE 5-1 Summary of Clinical Presentation and Location of Lesion in the Temporo-Occipital Vision Disorders
| Condition | Description | Location of Lesion |
|---|---|---|
| Dyschromatopsia | Inability to distinguish colors by hue | Inferior occipital lobe, occipitotemporal lobe |
| Color agnosia (and anomia) | Inability to name colors despite being able to match and sort items by color | |
| Alexia without agraphia or pure alexia | Inability to read despite preserved ability to write | Occipitotemporal lobes |
| Apperceptive (object) | Inability to recognize familiar objects despite intact visual acuity, contrast sensitivity, and often color perception and stereopsis | Bilateral occiptotemporal lobes |
| Associative agnosia | Sensory percept stripped of meaning | Bilateral occiptotemporal lobes |
| Prosopagnosia | Inability to recognize familiar faces | Bilateral (but right > left) occipitotemporal lobes |
| Environmental agnosia | Inability to recognize familiar environments | Right occipitotemporal lobes |
Agnosias are commonly subdivided on the basis of a distinction made by Lissauer,13 who posited in 1890 that the process of recognition has two distinct stages: apperception and association. Apperception is the ability to form a conscious percept of a sensory impression (e.g., an object), which can be thought of as the construction of different visual attributes of the stimulus into a whole percept. Patients who present with these difficulties are said to have the syndrome of apperceptive agnosia, sometimes referred to as apperceptive visual object agnosia.12 Association, on the other hand, refers to the imparting of meaning to the percept, achieved through the use of matching or linking the percept to a previous experience or knowledge.13 Patients with association difficulties, who are said to have the syndrome of associative visual agnosia, often have difficulty accessing the memory of an object’s name or its meaning from the visual stimulus, despite a more or less correctly perceived visual percept, and demonstrably intact knowledge (if accessed via other modalities) of the object’s name and semantic attributes.
Dyschromatopsia and Color Anomia
Dyschromatopsia is a rare acquired inability to discriminate colors by hue.14 Dyschromatopsia is most often associated with damage to the inferior part of the occipital lobes, in the fusiform gyrus. This corresponds to the area identified in functional experiments as the human color area (V4).15,16 Patients usually have difficulty in tasks such as sorting sets of colored counters. Hemidyschromatopsia may be more common; reflecting the location of damage, there is often an associated superior quadrantanopia on the same side. Color anomia is a disorder in which patients present with an inability to name colors but are still able to sort colored counters and differentiate between colors despite failure to name them.17
Alexia Without Agraphia
Alexia without agraphia (or pure alexia) is an acquired reading disorder in which the patient is unable to read, despite preservation of other aspects of language such as spelling and writing.18 Pure alexia is usually caused by an occlusion of distal (posterior) branches of the left posterior cerebral artery. The resultant damage is believed to interrupt the transfer of neural information from the visual cortex to the language cortex. Stroke is one of the most common causes of pure alexia, but more rarely it may result from closed head injury, tumor, occipital lobectomy, arteriovenous malformations, and Alzheimer’s disease. Further details are given in Chapter 3, Disorders of Language.
Associative Agnosia
Those patients with associative agnosia have apparently preserved sensory capability and may retain the ability to match and copy common objects but still fail to recognize them. Such cases led Teuber19 in 1968 to describe this as “a normal percept that has somehow been stripped of its meaning.” Some patients may have their problem limited to a specific class of objects, of which the best known is prosopagnosia—the failure to discriminate and recognize faces. Other object classes with which specific difficulties have been reported include animals or animal species, plants, foods, clothing, makes of cars, colors, or places. Some examples are discussed in the following paragraphs.
Associative Visual Agnosia for Familiar Faces: Prosopagnosia
Prosopagnosia is characterized by an inability to identify familiar faces. Patients may correctly identify the emotion but still fail to identify the face. Given that differences between faces are subtle, the patient with prosopagnosia has difficulty telling them apart but might be able to do so if there is a highly salient feature such as a birthmark or beard. The patient is often able to recognize familiar people from other attributes, such as gait, clothes, or voice. The particular attributes of faces and the task of their recognition are much debated in the literature, but it seems likely that face recognition requires some abilities that do not overlap with other visual recognition abilities. A number of studies have been made of the ability to recognize faces in the normal, upright orientation in a type of gestalt perception, compared with recognizing faces that are upside-down. The latter is a very much harder task for normal subjects and involves different (feature-by-feature rather than whole-object) visual processing. It is intriguing that some prosopagnosics do not have this face inversion effect and, paradoxically, may perform better with inverted faces, suggesting a specific deficit in whole-object processing.20
Prosopagnosia is usually caused by more anterior lesions in the inferior occipitotemporal cortex (the anterior aspects of the lingual and fusiform gyri).21,22 In most cases, bilateral damage is described, but some cases have occurred with just a right-sided lesion. This damage is most commonly a result of stroke, tumors, demyelination, or degenerative atrophy.
Many face recognition studies on normal subjects have been carried out using functional brain imaging techniques. Haxby and colleagues8 provided some of the key experiments, initially with positron emission topography and then with functional magnetic resonance imaging. Figure 5-5 is an example of the type of stimulus used for these experiments. Figure 5-6 shows the activation along the inferior occipitotemporal cortex in such a face-matching experiment.23
< div class='tao-gold-member'>

Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


