(1)
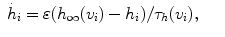
(2)
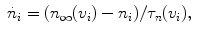
(3)
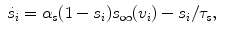
(4)
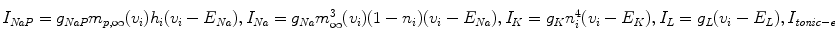 and
and ![$$I_{\mathit{CAN}} = g_{\mathit{CAN}}f([Ca]_{i})(v_{i} - E_{\mathit{Na}}).$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq2.gif)
The activation of the CAN current by the calcium concentration is given as ![$$f([Ca]_{i}) = (1 + (K_{\mathit{CAN}}/[Ca]_{i})^{n_{\mathit{CAN}}})^{-1}.$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq3.gif) The calcium dynamics is described as
The calcium dynamics is described as ![$$d[Ca]_{i}/dt = f_{m}(J_{ER_{\mathit{IN}}} - J_{ER_{\mathit{OUT}}})$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq4.gif) ,
, ![$$dl_{i}/dt = AK_{d}(1 - l_{i}) - A[Ca]_{i}l_{i}$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq5.gif) , in which
, in which ![$$J_{ER_{\mathit{IN}}} = (L_{IP_{3}} + P_{IP_{3}}[ \frac{IP_{3}[Ca]_{i}l_{i}} {(IP_{3}+K_{l})([Ca]_{i}+K_{a})}]^{3})([Ca]_{\mathit{ ER}} - [Ca]_{i})$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq6.gif) ,
, ![$$J_{ER_{\mathit{OUT}}} = V _{\mathit{SERCA}} \frac{[Ca]_{i}^{2}} {K_{\mathit{SERCA}}^{2}+[Ca]_{i}^{2}}$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq7.gif) and
and ![$$[Ca]_{\mathit{ER}} = \frac{[Ca]_{Tot}-[Ca]_{i}} {\sigma }.$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq8.gif) The meaning and values of other parameters are same as that in [10].
The meaning and values of other parameters are same as that in [10].
The calcium dynamics is described as , , in which , and The meaning and values of other parameters are same as that in [10].3 Bursting and Pattern Transition Mechanisms
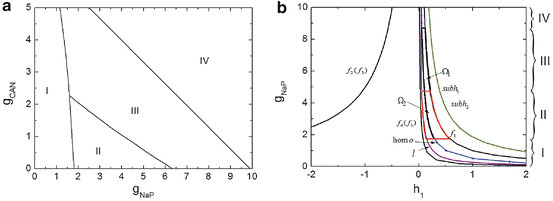
The activity patterns depending on parameters g NaP and g CAN are illustrated, as shown in Fig. 1a, The horizontal axis represents g NaP and the vertical g CAN . Two-coupled cells in the pre-BötC can generate two types of oscillations: the in-phase and anti-phase oscillations [1], so the two-parameter space can be divided into four regions: region I (silence), region II (in-phase bursting and anti-phase bursting), region III (in-phase bursting and anti-phase spiking) and region IV (in-phase spiking and anti-phase spiking). We chose g CAN = 0. 7 nS as a representative and explore the bursting transition mechanisms between these regions with g NaP changing. The systems yield a steady calcium concentration ![$$[Ca]_{1} = [Ca]_{2} = 0.02104057\,\mathrm{mV}$$](/wp-content/uploads/2016/09/A315578_1_En_49_Chapter_IEq9.gif) with
with 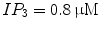 . h 1 and h 2 are almost equal when
. h 1 and h 2 are almost equal when 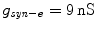 [1], we can consider h 1 and h 2 as one single slow variable, h 1. The two-parameter bifurcation analysis is shown in Fig. 1b. As g NaP increases, the systems undergo regions I, II, III and IV. The maximum and minimum values of limit cycles in the full system (1)–(4), we named “the slow variable regions” Ω 1 and Ω 2 [8], are appended in Fig. 1b. The bifurcation curves in regions Ω 1 and Ω 2 are different which play an important role in determining which types of bursting or spiking can occur.
[1], we can consider h 1 and h 2 as one single slow variable, h 1. The two-parameter bifurcation analysis is shown in Fig. 1b. As g NaP increases, the systems undergo regions I, II, III and IV. The maximum and minimum values of limit cycles in the full system (1)–(4), we named “the slow variable regions” Ω 1 and Ω 2 [8], are appended in Fig. 1b. The bifurcation curves in regions Ω 1 and Ω 2 are different which play an important role in determining which types of bursting or spiking can occur.
 Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
 Dynamic Temporal-Topological Structure of Brain Network Within ADHD
Dynamic Temporal-Topological Structure of Brain Network Within ADHD
 Activity Patterns in Cortical Dynamics and the Illusion of Localized Representations
Activity Patterns in Cortical Dynamics and the Illusion of Localized Representations
 on the Neural Energy Coding
on the Neural Energy Coding
 Cortical Singularities During the Cognitive Cycle Using Random Graph Theory
Cortical Singularities During the Cognitive Cycle Using Random Graph Theory
 as Bifurcations Shaped Through Sequential Learning
as Bifurcations Shaped Through Sequential Learning




with . h 1 and h 2 are almost equal when [1], we can consider h 1 and h 2 as one single slow variable, h 1. The two-parameter bifurcation analysis is shown in Fig. 1b. As g NaP increases, the systems undergo regions I, II, III and IV. The maximum and minimum values of limit cycles in the full system (1)–(4), we named “the slow variable regions” Ω 1 and Ω 2 [8], are appended in Fig. 1b. The bifurcation curves in regions Ω 1 and Ω 2 are different which play an important role in determining which types of bursting or spiking can occur.Related posts:
Integrated Neuropsychiatric Assessment System: A Generic Platform for Cognitive Neurodynamics Research
Dynamic Temporal-Topological Structure of Brain Network Within ADHD
Activity Patterns in Cortical Dynamics and the Illusion of Localized Representations
on the Neural Energy Coding
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
