(1)
Department of Behavioral Neurobiology, Institute of Experimental Medicine, Budapest, Hungary
Abstract
Understanding the neural control of a behavior necessitates the clear delimitation of the behavior that is controlled. While definitions on aggression are in no short supply, these often lack aspects that are crucial for this book: the cross-species validity of definitions, their corroboration with methods of measurement, and the clear delimitation of normal and abnormal forms. To address these issues, here we circumscribe aggressive behavior in two meanings, namely, in theoretical and practical terms. First, we will review intensional definitions that specify the necessary and sufficient conditions to differentiate this behavior from others. This will be followed by extensional definitions that list its main forms and subcategories. While these theoretical constructs clearly demarcate aggressive behavior, they cannot be directly used in research because they do not provide means to measure it. These means are provided by more practical—operational—approaches, particularly by the methodologies that are employed in aggression research to quantify aggressiveness, which will be corroborated with theoretical approaches.
The Latin original of the term aggression (“aggressio”) had a military connotation in ancient times and meant “undertaking a task” and “assault.” Its psychological and psychiatric meaning was acquired during the twentieth century; the definition “hostile or destructive behavior” was first recorded in 1912 in A. A. Brill’s translation of Freud.1 Hostile and destructive behavior, however, can be understood and measured in different ways especially when the term is used to denote behaviors expressed by different species.
Understanding the neural control of a behavior necessitates the clear delimitation of the behavior that is controlled. While definitions on aggression are in no short supply, these often lack aspects that are crucial for this book: the cross-species validity of definitions, their corroboration with methods of measurement, and the clear delimitation of normal and abnormal forms. To address these issues, here we circumscribe aggressive behavior in two meanings, namely, in theoretical and practical terms. Firstly, we will review intensional definitions that specify the necessary and sufficient conditions to differentiate this behavior from others. This will be followed by extensional definitions that list its main forms and subcategories. While these theoretical constructs clearly demarcate aggressive behavior, they cannot be directly used in research because they do not provide means to measure it. These means are provided by more practical—operational—approaches, particularly by the methodologies that are employed in aggression research to quantify aggressiveness. Animal and human methodologies will be evaluated comparatively; the boundaries between normal and abnormal aggressions will be in focus throughout. The last section of this chapter examines the area of validity of conclusions that are based on particular operational approaches and their relevance for understanding aggression control.
1.1 Demarcating and Characterizing Aggression: Theoretical Definitions
1.1.1 Basic Definitions
In principle, there are two types of definitions for “aggression”: one focuses on external appearance (i.e., the “form” of behavior), while the other focuses on its functions. “Behavioral form” or “phenomenological” definitions—highly preferred in the 1960s and 1970s—are usually restrictive but have the advantage of being simple and easily applicable to both animals and humans. In plain terms, aggressiveness can be defined as a behavior that delivers noxious stimuli to another organism (Buss 1961) or somewhat more explicitly: any behavior directed towards the goal of harm and injury (Baron 1977; Zillman 1979). These clear-cut definitions immediately raise a simple but relevant question: why should one organism deliver “noxious stimuli” to another? The explanation is provided by the other type of basic definitions, which attribute functions to this behavior. Functional definitions are likely based on the common observation that aggressiveness is present in any species that is endowed with the capacity of performing it, from invertebrates to humans.2 One cannot reasonably assume that aggressive behavior survived evolutionary pressures for hundreds of millions of years in so many species without providing some functional advantages. It is generally believed that aggression is the most efficient form of competition that bridges the gap between the limitedness of resources on one side and the individual needs of survival and the more general “evolutionary need” of leaving as many descendants as possible on the other side. Thus, the proximate function of aggression is resource competition that ensures survival, while the ultimate function is reproduction that ensures contribution to the gene pool of the next generation. This dual goal is achieved by fights over food, territory, mating partners, etc., and by protecting offspring by aggressive means. This utilitarian view on aggression was formulated very early (Darwin 1871) and was maintained ever since (see Archer 2009; Lindenfors and Tullberg 2011; Weiger and Bear 1988 as examples).
Thus, individuals harm each other to get access to resources and to ensure gene transmission over generations in a world where resources are limited and the delivery of harm is more efficient than negotiation.3 At the first sight, the functional definition applies to animals only, because survival and the ability to leave progeny rarely depend on aggression in humans. Human societies consist of highly interdependent individuals, which are expected to negotiate rather than fight for resources. Still, human aggression is often gain oriented; this type of human behavior is commonly called instrumental aggression (Feshbach 1971). Moreover, particular forms of aggression (usually considered legitimate) serve survival in the form of self-defense, the defense of offspring, or the defense of others (Archer 2009; Björkqvist et al. 2000). Furthermore, intricate forms, e.g., relational and indirect aggressions, are means of social competition that are employed at a wide scale in human societies. Because human aggression may become a matter of survival and inclusive fitness under certain conditions, one should attribute functionality to human aggression as well. Taken together, these considerations show that (1) aggression is omnipresent in arthropods and vertebrates and is shown by all lower animals that are endowed with the capacity of performing it and (2) it confers competitive advantage in animals and has similar roles in people under particular conditions.
A final clarification that intensional definitions need refers to the word “harm.” In his influential work on aggression, Lorez (1963) notes that “crude” forms of aggression (i.e., the delivery of injuries) gradually progress into “symbolic” actions that fulfill the function of communication. The reason is that aggressiveness involves considerable risks for both the recipient and perpetrator. Either of the fighting parties can suffer injuries, and both lose energy reserves that may become vital in a subsequent challenge (e.g., predator attack) (Haller 1995). Therefore, injurious behaviors not only are gradually replaced with but are also preceded by less risky behaviors, e.g., threat signals that offer the chance of withdrawal before the actual fight is started. Such forms of behavior are integrant parts of the aggressive repertoire of animals (see van Staaden et al. 2011 for a review). Human aggression also entails threats, e.g., threatening facial expressions and body language, verbal warnings, etc. It is generally considered that threatening with physical harm is an aggressive act per se. On the other hand, harm should not always be understood in a physical sense. Manipulating social environments (e.g., by malicious gossiping, ignoring, and coercing) may hurt a person in a nondirect manner and may ultimately disrupt his/her social status (Bjorkqvist and Niemela 1992; Coccaro et al. 1997a, b; Galen and Underwood 1997; Gomes 2007; Paquette and Underwood 1999). Social manipulation is not uncommon in animals; the main behavioral tool is social exclusion in their case (see Sasaki and Uchida (2012) for a review and Goodall (1986) for studies in chimpanzees). Therefore, a more complete definition should make reference not only to harm and injury but also to threatening with these; in addition, it should consider “harm” in both physical and psychological terms (Björkqvist et al. 2000; Brower and Price 2001). Further specifications of the word “harm” include intentionality (to exclude accidental events) and exclude benefits for the victim (Berkowitz 1993, 1994; Björkqvist et al. 2000; Bushman and Anderson 2001; Feshbach 1971). As such, aggression includes neither accidental harm nor harm inflicted for positive ends (e.g., to discourage social parasites, to discipline offspring, or to maintain cooperative behavior; Clutton-Brock and Parker 1995). To avoid an impossibly complex sentence, we define aggression below by pointing out its essence and adding modifiers.
Aggression is a behavioral means of resource competition that is based on the delivery of or threatening with harm. Harm should be understood in both physical and psychological terms, should be intentional, and should benefit the victim neither directly nor indirectly.
1.1.2 Definitions for Subtypes
The definition adopted above shows that aggression is not a unitary phenomenon and cannot be the object of studies in such general terms. Therefore, aggression was divided into several subtypes along a variety of classification principles. The form of aggression is one such principle; in humans, subtypes are called physical, verbal, and indirect aggressions (Bjorkqvist and Niemela 1992; Gomes 2007; Paquette and Underwood 1999; Vaillancourt and Sunderani 2011). Physical aggression involves physical harm; verbal aggression (including body language) covers threats that aim at intimidating partners, while indirect aggression consists of social manipulation that inflicts psychological harm in most cases. In some papers, indirect aggression was restricted to or included aggression towards objects (Coccaro et al. 1997a, b; Crowe 1974). This approach was considered outdated by papers published later (Gomes 2007). We suggest here that aggression against objects (e.g., slamming doors) should be considered part of “verbal” aggression when its aim is intimidation and displaced aggression when it serves to channel off emotions. This issue will be detailed below. Rodent analogs of these behavioral categories include biting attacks (physical aggression), offensive threats (verbal aggression), and social exclusion (indirect aggression). Phenomenologically delimited subtypes can be graded according to their precise forms and the severity of consequences. For example, fighting with bare hands is considered milder than weapon use; injuring is a lesser form of aggression than killing (Björkqvist et al. 2000; Holm 1982). Similarly, body language and verbal threats may be perceived as graded versions of “verbal” aggression. Finally, indirect aggression can be divided into direct (conspicuous) and hidden forms (Gomes 2007).
The reason and emotional background of aggression can also serve as criteria for typifying this behavior. Emotional and instrumental forms are the most widely used categories (Blair 2001; Feshbach 1971; Kempes et al. 2005; Lopez-Duran et al. 2009; Raine et al. 2006; van Bokhoven et al. 2005; van Honk et al. 2010). The former (often called reactive) consists of uncontrolled aggressive outbursts that are elicited by perceived threat or provocation. This type of aggression is characterized by high levels of bodily arousal. Instrumental aggression (also called proactive) is controlled and goal directed. It is usually associated with low levels of arousal. Emotional and instrumental aggressions are associated with different types of criminal aggression and aggression-related psychopathologies, as discussed below. There are several rodent models of abnormal aggression, where important aspects of emotional (reactive) and instrumental (proactive) aggression were mimicked (see below). A specific form of emotional responses is displaced aggression; this behavior is directed towards a third party under conditions of frustration and distress. While this behavior is commonly studied in people (Denson et al. 2006; Fitz 1976; Kaufmann and Feshbach 1963; Pedersen et al. 2000; Reijntjes et al. 2007), similar studies in animals are sparse although displaced aggression is readily shown by vertebrates from fish to rats (Overli et al. 2004; Nephew et al. 2003; Hoffmann et al. 1987).
Other frequently used categories are overt and covert aggressions. These may be perceived and defined in different ways. Popma et al. (2007a), for instance, use the phrase “overt aggression” as a synonym of physical and verbal aggressions combined, while “covert aggression” depicts emotional components in their terminology (e.g., hostility, angriness, jealousy, etc.). In other papers, “overt aggression” includes physical fights, weapon use, cruelty, robbery, and sexual abuse, while “covert aggression” is defined as housebreaking, steeling, fire setting, lying, truancy from home, etc. (van Bokhoven et al. 2005). The phrases “overt” and “covert” are sometimes perceived as synonyms of physical and indirect aggressions, respectively (Vaillancourt and Sunderani 2011).
The brief review of various classifications shows that the terms are not used consistently, and there are overlaps between various categories. For example, indirect aggression is instrumental in many cases; covert aggression is considered synonymous with hostility and anger according to one categorization and with nonviolent lawbreaking according to the other. Any combination is possible between emotional (reactive) and instrumental (proactive) aggressions on one side and physical and verbal aggressions on the other. In addition, such subcategories do not categorize perpetrators but actions: one and the same person may show emotional aggression under certain circumstances and instrumental aggression under others. Despite inconsistencies and overlaps, these concepts are very useful for differentiating and studying distinct classes of behaviors.
Another criterion that may be used to typify aggression relates to its normality. Given the importance of this criterion for the present work, it will be presented in a separate section.
1.1.3 Normal and Abnormal Aggressions: Basic Principles
As shown above, aggression is ubiquitous in animals where it plays important functions. For this reason, abnormality may be perceived unlikely as far as animals are concerned. On the other hand, the high level of socialization and the strong moral, legal, and social stigmatization of aggressiveness may render aggression per se abnormal in humans. In simple terms, one may be tempted to believe that aggression is always normal in animals and is always abnormal in people. Neither assumption seems true.
Conflicts in animals are settled according to certain rules, which result from evolutionary pressures against dangerous forms of competitiveness (Hinde 1974). Intraspecific aggression is not aimed at inflicting lethal injuries but at ensuring access to resources. Species would be rapidly eliminated if its individuals risked their lives to obtain food, mating partners, etc. Moreover, the near-emptying of biochemical energy stores by excessive fighting per se may put the life of animals at risk (Haller 1995). Evolution shaped aggression such that these risks are avoided. Aggressive intent is signaled in advance to allow withdrawal, the level of aggressiveness is kept between certain limits to minimize risks and excessive energy losses, attacks are targeted towards body parts that lack vital organs and are not excessively exposed to infections, aggressiveness is rapidly deferred by submission or flight by the opponent, and, finally, females (rate-limiting factors in reproduction) are largely protected from attacks by males (not considering sexual coercive acts). Deviations from these rules capsize the balance between costs and benefits, render aggression nonfunctional, and provide a handle for differentiating normal and abnormal aggressions in animals (Haller et al. 2001, 2005a; Haller and Kruk 2006). Thus, abnormal aggression is defined in terms of natural rule breaking and the lack of functionality in animals.
Human aggression is considered abnormal if it violates moral and legal rules or if is associated with psychopathologic conditions. These two principles (morality, law and mental health) are closely related. An interesting study by Björkqvist et al. (2000) demonstrated that people base their attribution of aggression on factors that are consistent with the severity of punishment in sentences for different crimes of violence; thus, moral and legal thinking is closely related as far as aggression is concerned. Significant overlaps were also found between the behavioral and legal correlates of aggression as well as between aggression-related psychopathological constructs and criminal behavior (Coccaro and McNamee 1998; Lee and Coccaro 2001; Gao and Raine 2010; Yang et al. 2008). Moreover, the characterization of aggression-related psychopathologies by DSM-IV-TR explicitly includes rule breaking among the symptoms of conduct, oppositional-defiant, and antisocial personality disorders and implicitly in the case of others (e.g., intermittent explosive disorder) (American Psychiatric Association 2000). Overall, this source considers psychological conditions pathological if they inflict suffering; cause significant impairment in social, occupational, or other important areas of functioning; and are persistent. Persistent violent criminal behavior fulfills these criteria.
In legal terms, the key factor that differentiates normal and abnormal aggressions is rule breaking. For example, violence was recently defined as the infliction of physical harm “in violation of social norms” (Brower and Price 2001). In psychopathological terms, aggression is abnormal if it makes the individual dysfunctional. Persistent aggressiveness leads to social exclusion, to the loss of jobs, to destruction of families, etc.—briefly, it imperils social, occupational, and other important areas of functioning. Thus, legal and psychopathological delimitations of human abnormal aggression, when taken together, replicate the definition of abnormal aggression proposed above for animals; human abnormal aggression can be defined in terms of rule breaking and dysfunctionality. Differences between animals and humans lie in details only. For example, rules are shaped by evolutionary pressures in animals, while human rules are formalized. The nature of punishment may also be dissimilar; abnormally aggressive animals may take punishment in the form of defeats when meeting opponents of superior fighting abilities, while humans risk the same and legal punishment in addition. Even the very long-term consequences appear comparable; excessive increases in the share of abnormally aggressive individuals imperil the success of animal species on an evolutionary scale; in the case of humans, the same jeopardizes the success of societies on a historical scale. Given these close parallels, we define below abnormal aggression by summarizing all the considerations outlined above.
Abnormal aggression breaks natural or formalized rules and is dysfunctional on the long run in terms of negative consequences for the perpetrator. This definition implies that certain forms of aggression are not abnormal in humans. Indirect aggression, for instance, does not break the law and may reach its function if carried out skillfully (i.e., it may confer competitive advantage). The same holds true for verbal aggression in many instances. Even physical aggression may be normal if performed by authorized bodies (e.g., law enforcement) or if lawful (e.g., self-defense). On the other side, some forms of aggression may be abnormal in animals. In the laboratory, such abnormal forms can be induced by treatments that mimic etiological factors of human abnormal aggression. Although the issue was not investigated systematically, the conditions leading to abnormal aggression (e.g., excessive stressors suffered in certain periods of life; see below) are likely occurring under natural conditions, which makes it probable that abnormal aggression exists in free-ranging populations.
1.1.4 A Theoretical Classification of Aggressive Behaviors
We suggest here a classification of aggressive acts based on the constructs overviewed above. The classification was visualized in Fig. 1.1. This figure will be used as a template for evaluating the correspondences between theoretical definitions and operational approaches. Animal aggression was not considered at this stage, but will be included in the system later on. We believe that there are three principles along which aggression should be categorized: behavioral form, reason, and abnormality.
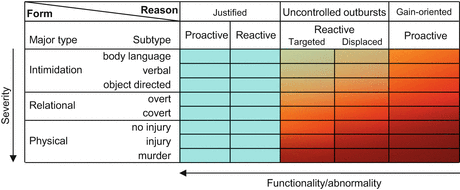
Fig. 1.1
The classification of aggressive acts based on behavioral characteristics (row headings), type/reason (column headings), and the level of functionality/abnormality (represented by color code; functionality decreases from blue to dark red). For details see Sect. 1.1.4
The forms of behavior were listed in the rows of the figure. The three main categories mentioned above (physical, verbal, and indirect) were slightly changed. Particularly, the phrase “verbal aggression” was replaced with “intimidation” to accommodate nonverbal forms that have the same function: body language and object-directed aggression when the latter is displayed with the aim of intimidation (e.g., hammering the table, slamming doors, etc.). Other forms of object-directed aggression (e.g., breaking objects to channel emotions) are considered displaced aggression. Instead of the word “indirect aggression,” we use here “relational aggression” to avoid the inconsistencies mentioned above; the two phrases are often used as synonyms in the literature. The severity of the behaviors increases downwards; intimidation is considered the least severe form because it only threatens with harm; relational aggression involves harm in psychological meaning, while physical aggression in physical meaning. These major forms were also graded according to more specific behavioral aspects. Body language was considered a milder form of intimidation than verbal aggression or object-directed aggression. In the case of indirect aggression, overt forms were considered less severe because they allow for mitigation. The level of injury infliction differentiates physical forms of aggression.
The reasons of being aggressive were listed in column headings. The following reasons were differentiated: legitimate uses of aggression (e.g., the defense of others and self-defense), the inability to control emotions and behavior (uncontrolled outbursts of anger), and gain-motivated aggression. Legitimate uses of aggression were further subdivided into proactive and reactive forms. The former is typical to law-enforcement bodies, while the latter is usually shown by those defending themselves from attack. Uncontrolled outbursts of anger were further subdivided into target oriented (e.g., aimed at the source of threat or provocation) and displaced (channeled towards objects or a third party). The legitimacy of the three main reasons decreases from left to right. Any form of defense is considered legitimate provided that it is proportional; disproportional aggression belongs to one of the subsequent two categories. Uncontrolled outbursts of anger cover aggressive acts that are not justified by the situation (i.e., they are out of proportions). This form of aggression is usually called reactive/emotional and is characterized by strong emotional reactions. Such actions are not legitimate, even less so if the target is a third party (i.e., displaced aggression against people). Yet in legal and moral terms, acting under psychological distress is extenuative to a certain extent; therefore, reactive/emotional aggression was considered less rule breaking (somewhat more legitimate) than instrumental aggression. The least legitimate forms of aggression are those performed for gain.
The color code of Fig. 1.1 denotes abnormality based on the functionality of behavioral acts. As aggressiveness is believed to be a means of competition, functionality was evaluated from the point of view of competitive advantages. On the short term, any of the aggressive behaviors listed in Fig. 1.1 may be considered functional according to this criterion. However, certain types of aggressive behaviors are competitively disadvantageous on the long run, for which these were classified as dysfunctional. For example, excessive and uncontrolled outbursts of aggression disrupt families, social networks, and work places, which renders this behavior dysfunctional overall, even if such outbursts may serve momentary interests in terms of assertiveness under particular circumstances. Similarly, instrumental aggression may make the individual successful in certain competitive situations, but on the long run, they involve personal risks and social punitive reactions that make instrumental aggression dysfunctional overall. By contrast, defense-oriented aggression is advantageous on the short run (it averts dangers) and does not entail long-term risks in legal or other terms. Therefore, defensive aggression was considered functional. Social acceptability and the severity of aggressive acts were also taken into account to grade the dysfunctional nature of aggressive behavior. Intimidation and relational aggression are ubiquitously used by humans as means of competition. Expressing displeasure by body language, verbally, or by object-directed aggression (e.g., slamming doors, hammering tables) is quite common during conflict. Spreading rumors, excluding people from social networks, or setting confederates against competitors is similarly common within social groups. Regular involvement in such behaviors may disfavor the individual on the long run, but negative consequences are not as serious as with physical forms of aggression. Therefore, these behaviors were considered less dysfunctional.
1.2 Quantifying Aggression: Operational Approaches
The definitions discussed above are intended to demarcate aggressive from other types of behavior and to separate its subtypes. Neither definition is sufficient per se to study aggression; these require specific methodologies. This chapter describes the main approaches in animals and humans, with a special focus on quantitative measures and the relevance and usefulness of the findings obtained for understanding normal and abnormal aggressions. We mention that each model and approach will be discussed in more detail in the subsequent chapters. Here we provide a general overview only.
1.2.1 Models of Normal Aggression in Animals
In animals, aggression is usually studied “in action,” i.e., while aggressive acts are actually performed. This is in sharp contrast with the most frequently employed approaches in human aggression research (see below). Even in animals, however, the neurobiology of aggression is rarely studied under entirely naturalistic conditions, partly because the required equipment and the data sampling procedures need laboratory environments, partly because laboratory studies ensure a stability of conditions that are rarely met in nature. Therefore, aggression is usually studied by employing models. The most fruitful models used so far are brain-stimulation-evoked aggression models and the resident-intruder test (the latter including studies on maternal aggression). Other methodologies include defensive, fear-induced, shock-induced, and social aggressions. Models of predatory aggression also exist; these will be discussed later on in relation with intraspecific aggression only. The methodologies discussed here are assumed to address various aspects of normal aggressiveness, i.e., are supposed to model aggressiveness shown by animals under naturalistic conditions. We will briefly characterize the main models below.
1.2.1.1 The Resident-Intruder Test in Males and Females
In its “classical” version, male subjects (usually rodents) are moved to an individual cage and are faced with an opponent (intruder) after a suitable time, usually after a couple of days. The behavior of rodents in this model was first described by Grant and Mackintosh (1963) and can be considered the most popular aggression model ever since. It is believed that individual housing activates territoriality; as such, aggressiveness elicited by this test is perfectly in line with functional definitions of aggression, because territories are the resources animals are most frequently fighting for under natural circumstances. Aggressiveness is increased by cohabitation with a female; this condition may add to the functional relevance of the model, because reproduction-related aggressiveness is also among the strongest naturalistic motivations. In phenomenological terms, aggressiveness shown in this model consists of various social, offensive, defensive, and dominance-related postures as well as biting attacks. The latter are overwhelmingly targeted towards the back and flanks of opponents (i.e., regions that contain no vital organs and are relatively less prone to infections); in addition, the frequency of biting attacks is reasonable (5–10 bites over a usual test). Thus, the behavior of subjects follows the “rules” outlined above. The latency of attack, the number of biting attacks, and the duration of aggressive postures (threats) are used as measures of aggressiveness. Social behaviors may be studied to complete the picture, while the durations of defensive and dominance-related behaviors may be used to evaluate the outcome of the fight. The contribution of this model to aggression research is substantial and diverse; it was extensively used to elucidate the roles of environmental conditions and hormones; it was used in pharmacology-oriented studies and those focusing on genetic factors, as well as in studies on mechanisms of neurobiological control. The models of abnormal aggression are also based on the resident-intruder test.
Albeit regular resident-intruder tests can be performed in females, aggressiveness is rather low in this gender for which this approach is rarely employed. By contrast, females violently defend pups from unfamiliar males that are prone to kill them. The behavior differentiates targets; female intruders (less dangerous for pups) are rarely attacked. This behavior called maternal aggression is intensely studied and can be considered the “female variant” of the resident-intruder test. The first recorded use of the model dates back to the early 1970s (Gandelman 1972). While aggressiveness shown in the resident-intruder and maternal aggression models is similar in many respects, there are important differences. In contrast to males, females are less tending to signal their attacks by offensive threats and readily attack vulnerable body parts of intruding males (Parmigiani et al. 1988). Albeit this behavioral profile is akin to the one marked as abnormal in males, it seems perfectly functional in this case, because subjects fight opponents that are superior in body size and defend resources of utmost value. An interesting feature of maternal aggression is its association with reduced stress responses that is also seen in certain models of abnormal aggression (see below). Particularly, maternal aggression is associated with low glucocorticoid stress responses (Neumann 2001; Neumann et al. 2001); moreover, acute stressors suffered before the encounter and increased stress responsiveness (as an individual feature) decrease maternal aggression (Gammie and Stevenson 2006; Gammie et al. 2005). A recent study in humans also showed that reduced autonomic arousal disinhibits female aggression in humans (Hahn-Holbrook et al. 2011). Taken together, these findings suggest that maternal aggression—although normal—shows similarities with certain models of abnormal aggression in both behavioral and emotional terms; as such, the maternal aggression paradigm offers unique possibilities to understand the control of aggression.
1.2.1.2 Brain-Stimulation-Evoked Aggression
Although this is not a naturalistic model of aggression, it is believed to be based on the activation of brain mechanisms that underlie natural forms of aggressiveness. As such, this induced behavior may reveal the neurobiological mechanisms of aggression in general; this justifies its presentation in this section. The methodology was introduced by Hess (1928), who observed that the electrical stimulation of particular hypothalamic sites rapidly induces biting attacks directed towards social partners. In the lack of the latter, no behavioral responses are evident. This methodology contributed to aggression research by delimitating aggression-related hypothalamic brain regions and by elucidating the major components of aggression-related neural networks (for reviews see Kruk 1991; Siegel et al. 1999). The measure of aggression is the strength of the electrical stimulation that is required to elicit aggression, which is inversely related to aggressiveness (larger current thresholds denote suppressed aggression). Networks subserving attacks were usually identified by the parallel stimulation of two or more brain areas and by the combination of stimulation and lesion techniques. The technology is also suitable for studies on the pharmacology of aggression; in this case, stimulation is combined with the systemic or local brain application of pharmacological agents. Electrical stimulation is rarely used today to study aggression, perhaps because it requires sophisticated equipment and deep knowledge in the field of physics. Another reason may be its low neuroanatomical resolution because electric currents stimulate all the neuron and at least some of the passing fibers that surround the electrode. The place of this technology may be overtaken by optogenetics which has a considerably better neuroanatomical specificity (Lin et al. 2011). It is important to note that brain-stimulation-evoked aggression fulfills most criteria based on which normal and abnormal aggressions are differentiated. Vulnerable targets, females, submissive animals, etc., are readily attacked, and the behavior is not functional by any means, because it is motivated by stimulation, not by the social situation. Nevertheless, it reveals the basic mechanisms of aggression, and the neural networks identified with this methodology guided research made in more naturalistic models.
1.2.1.3 Shock-Induced Aggression
It was observed rather early that the administration of mild electric shocks immediately elicits aggressiveness in a novel environment where spontaneous aggression is rare (Eichelman 1971; Powell et al. 1969). Albeit the model gained some popularity in the 1970s and 1980s and is still in use today, it did not have an impact comparable to the two models discussed above. A PubMed search with the search terms (rat [title/abstract] OR rats [title/abstract] OR mouse [title/abstract] OR mice [title/abstract] AND aggression [title/abstract] AND shock-induced [title/abstract]) returned 76 hits, while the same search term without the phrase “shock-induced” returned over 2,300, suggesting that the model is not used very frequently. The likely reason is the strange behaviors elicited by shocks. Unfortunately, the studies employing the model tend to provide global scores for aggressiveness, for which it is difficult to evaluate what actually happened during the test. However, “aggressiveness” elicited by shocks is a mixture of offense and defense according to our personal experience. Biting attacks are rare.
1.2.1.4 Fear-Induced Aggression
Models of this type involve the exposure of subjects to conspecifics, predators, or humans under conditions that elicit fear (conspecifics: Blanchard and Blanchard 1989; predators: Blanchard et al. 1990; humans: Plyusnina and Oskina 1997). The models in use are centered on the notion of defensive aggression, supposed to be different from offensive aggression. Such models are used by a restricted number of research groups only. Reasons may be multiple and may range from a faulty understanding of the issue by the aggression research community to practical and theoretical problems associated with such models. To name a few of these: (1) offense and defense are always intermixed irrespective to the status of the individual; (e.g., both residents and intruders show offense and defense in the resident-intruder test), (2) situational and behavioral defensiveness and offensiveness are also intermixed (e.g., residents defend their territory by offensive behaviors), and (3) strong defensive responses (e.g., those elicited by predators) are so much mixed with fear that they were proposed as models for various types of anxiety (panic: Blanchard et al. 1997; post-traumatic stress disorder: Hammack et al. 2012). While defensive behaviors are just as important for the individual as offensive ones, and defense may involve acts of offense under particular conditions, they seem to be motivated primarily by fear. As such, these behaviors are only partly consistent with phenomenological definitions and inconsistent with functional ones, for which will receive marginal attention in this book.
1.2.1.5 Social Aggression
The closest analogs of human aggressive behavior are tests where subjects live in colonies, and aggressiveness is studied in conjunction with their social life. The prototypical example of such models is the visible burrow system developed by Blanchard et al. (1988). In this model, rat colonies live together for long periods of time in large enclosures that include burrows (natural living spaces for rodents). Less ambitious models also exist; in their case, the length of cohabitation is restricted to a couple of weeks and the enclosure is smaller and does not include burrows (Mikics et al. 2007; Millard and Gentsch 2006). Colony-based models were used to evaluate the impact of group living on the ontogenetic development of emotional and stress responses (Kaiser et al. 2007) and to study social integration in animals submitted to models of abnormal aggression (Tulogdi et al. 2014). Despite their obvious value for studying aggression in social species (e.g., rats), these models are rarely employed, likely because they are extremely labor intensive and require large laboratory space.
1.2.2 Models of Abnormal Aggression in Animals
The idea of studying abnormal forms of aggression in rodents is relatively new in aggression research: our lab was the first to propose a model (Haller et al. 2001). This does definitely not mean that abnormal aggression was not seen in rodents before this publication; moreover, models used earlier may readily be included into the large family of abnormal aggression models. There were three concepts that differentiated this publication from the earlier ones. First, we deliberately generated the model by making use of data on the etiological factors of antisocial aggressiveness. Second, we detached from the generally employed quantitative approach: instead of counting attacks, we focused on their form and behavioral context. Third, attention was not restricted to aggressive behavior per se; we sought to investigate other features related to antisocial aggressiveness (e.g., autonomic responses, anxiety, and social behaviors beyond aggressiveness) (Haller et al. 2004). Based on the principles underlying this experimental work, we developed a general concept for abnormal aggression models (Haller et al. 2005a; Haller and Kruk 2006). The same complex approach was employed when our second model of abnormal aggression was developed (Toth et al. 2008, 2011).
Models of abnormal aggression gained terrain in two directions: models used earlier were reevaluated in the light of this concept (see Miczek et al. (2013) for a review), and new models were developed to address other etiological factors of abnormal aggression. The criteria that differentiate normal from abnormal aggression were reformulated in several ways by the authors who undertook research in this relatively new field. Nevertheless, the essential points remained the same. Rats are submitted to procedures that mimic etiological factors of aggression-related psychopathologies and are tested for abnormal attack features in the resident-intruder test. These features can be of three types: mismatch between provocation and response, disregard of species-specific rules, and insensitivity towards the social signals of opponents.
The etiological factors considered in abnormal aggression models include the application of stressors at various ages, treatments with drugs, and genetic manipulations. We will review below the main models based on the type of the etiological factor that produced abnormal aggression. Two models will be discussed separately, because of their specific features. One model was based on the repeated exposure of rats to victories, while the other on the suppression of glucocorticoid production. In all cases, the emotional background of aggression will be evaluated, because this feature seems relevant to the type of aggression performed and the classification of the models.
1.2.2.1 Stress Models
Human studies show that stressors suffered in early childhood or adolescence (e.g., abuse, maternal and social neglect, as well as other types of severe stressful life events) considerably increase the risks of violent and antisocial behaviors later in life (Attar et al. 1994; Chapple et al. 2005; Dodge et al. 1990; Loeber and Hay 1997; Paperny and Deisher 1983; Trentacosta and Shaw 2008; Widom 1989; Windle and Windle 1995). In addition, acute provocations elicit immediate increases of aggressiveness in adulthood (Baldwin and Randolph 1982; Pedersen et al. 2000; Archer and Benson 2008).
These findings served to develop a series of abnormal aggression models. Repeated maternal separation was used to mimic early maternal neglect (rats: Veenema et al. 2006), postweaning social isolation was used to model early social neglect (mice: Bibancos et al. 2007; rats: Toth et al. 2008; Wall et al. 2012 4), early social subjugation was used to model adolescent abuse (hamsters: Delville et al. 1998; rats: Cunningham and McGinnis 2008), and, more recently, strong physical stressors were repeatedly administered to pubertal rats to mimic the impact of nonsocial stressors suffered at this age (Márquez et al. 2013). In these models, subjects were exposed to the stressors in the life periods indicated above, while aggression was studied by the resident-intruder test. Finally, the immediate effects of stressors were mimicked by the “instigation” model of aggression, where mice are exposed to territory intrusion without being able to contact the opponent physically, which is followed by exposure to a second opponent which now is physically accessible (Fish et al. 1999). Noteworthy, models were attributed to authors who were the first to provide data from which abnormal features of aggression could be deduced.
The models listed here are in different stages regarding the characterization of aggressive behaviors that result from stress application. Only disproportionality (excessive attack counts) denotes abnormality in the maternal separation (Veenema et al. 2006) and instigation models (Fish et al. 1999). Increases are dramatic in the latter, which renders this model appropriate for investigating the neurobiological mechanisms underlying the immediate effects of stressors on abnormal forms of aggression. In the maternal separation model, dramatic increases in aggression were noticed in juveniles but not in adults, suggesting that the consequences of this stressor are either washed out during development or that adult behavior needs more detailed analysis (Veenema et al. 2006; Veenema and Neumann 2009). Note that abnormal attack features may be present even in the absence of increased attack counts (see below). Interestingly, maternal separation decreased aggression in mice (Tsuda et al. 2011); unfortunately, however, aggressive behavior was not presented in detail in this paper. Dramatic increases in aggression were noticed in the early subjugation model, where, in addition to this quantitative change, an interesting phenomenon also occurred: while aggressiveness increased when subjects faced smaller opponents, it markedly decreased when opponents faced similar or larger opponents (Delville et al. 1998; Ferris 2003). We suggested that this “offensive ambiguity” may be listed among abnormal attack features (Haller 2013). In rats, early subjugation also increased aggressiveness; this was evident when rats were provoked during aggressive encounters by regularly applied tail pinches (Cunningham and McGinnis 2008). Based on the composite score of aggressiveness provided, the increase was dramatic (four–fivefold). Aggression towards females was not increased in the rat subjugation model (Frahm et al. 2011). Taken together, these data suggest that early subjugation in rats can also be considered a model of abnormal aggression based on the etiological factor and the exacerbated response to provocation.
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


