Smell and Tast
All neurons in the brain respond to chemical transmitters, so chemosensitivity is hardly a specialization of function at all. Here we shall be considering only chemical stimuli that originate outside the body, with olfaction (smell) and gustation (taste). The functions of the chemoreceptors that monitor the composition of the blood, and are used in the regulation of autonomic and hormonal functions, are discussed in Chapter 14.
Olfaction
Receptors
If you have had the privilege of dissecting a human head you will be aware that the nose has an internal complexity that is startling in comparison with its rather drab exterior.  The surface area of the nasal cavity is enormously inflated by the presence of three conchae on each side, highly vascular organs covered with erectile tissue whose function is primarily to moisten and warm the incoming air, and conversely to limit the loss of heat and water in the air that is expired. The olfactory receptors form part of the olfactory epithelium, tucked away in the olfactory cleft right at the top of the cavity, and in normal quiet breathing only a very small proportion of the air actually reaches the olfactory epithelium.
The surface area of the nasal cavity is enormously inflated by the presence of three conchae on each side, highly vascular organs covered with erectile tissue whose function is primarily to moisten and warm the incoming air, and conversely to limit the loss of heat and water in the air that is expired. The olfactory receptors form part of the olfactory epithelium, tucked away in the olfactory cleft right at the top of the cavity, and in normal quiet breathing only a very small proportion of the air actually reaches the olfactory epithelium.  But when we sniff, turbulences are set up round the conchae, and an appreciable fraction of the air gets to the olfactory receptors. This fraction is critically dependent on the state of the conchae: if you have a cold, they tend to become engorged with blood, hindering the passage of air to the higher regions and causing the familiar partial loss of smell.
But when we sniff, turbulences are set up round the conchae, and an appreciable fraction of the air gets to the olfactory receptors. This fraction is critically dependent on the state of the conchae: if you have a cold, they tend to become engorged with blood, hindering the passage of air to the higher regions and causing the familiar partial loss of smell.
The surface area of the nasal cavity is enormously inflated by the presence of three conchae on each side, highly vascular organs covered with erectile tissue whose function is primarily to moisten and warm the incoming air, and conversely to limit the loss of heat and water in the air that is expired. The olfactory receptors form part of the olfactory epithelium, tucked away in the olfactory cleft right at the top of the cavity, and in normal quiet breathing only a very small proportion of the air actually reaches the olfactory epithelium. But when we sniff, turbulences are set up round the conchae, and an appreciable fraction of the air gets to the olfactory receptors. This fraction is critically dependent on the state of the conchae: if you have a cold, they tend to become engorged with blood, hindering the passage of air to the higher regions and causing the familiar partial loss of smell.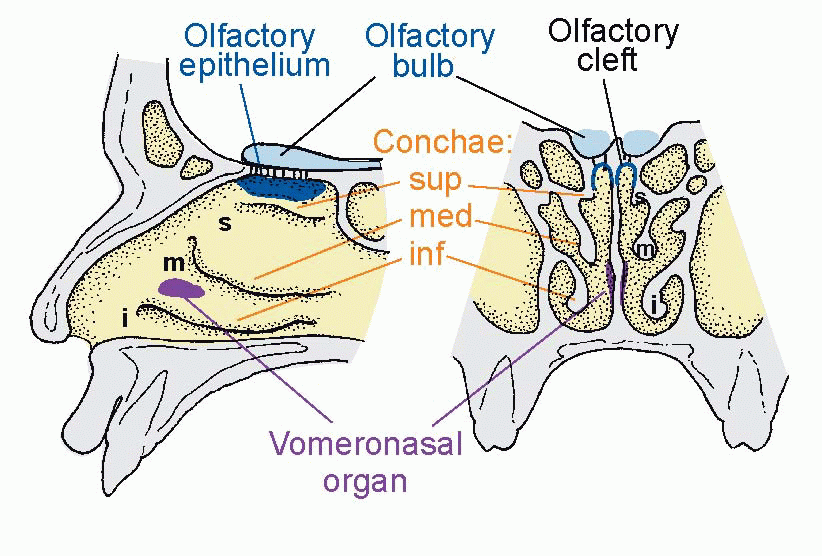 |
Humans are microsmatic animals: smell plays a far smaller part in their sensory world and in the regulation of their actions than in the case of macrosmatic animals such as the dog, and their olfactory sensitivity is in general correspondingly reduced. This is reflected in the small area of our olfactory epithelium: some 10 cm2 in all, about half that of the cat, despite its very much smaller head. This epithelium has a number of easily recognizable features. It contains Bowman’s glands, producing a lipid-rich secretion that bathes the surface of the receptors; this means that a substance must to some extent be lipid-soluble to have an odour. Another characteristic is that one of the types of epithelial cells contains granules of pigment: the depth of colour in different species is often correlated with olfactory sensitivity, being light yellow in Man and dark yellow or brown in dogs. It has sometimes been suggested that this pigment may play some part in the mechanism of olfactory transduction, perhaps being involved in the absorption of some kind of radiation such as infra-red, an attractive but controversial idea discussed later (p. 181).
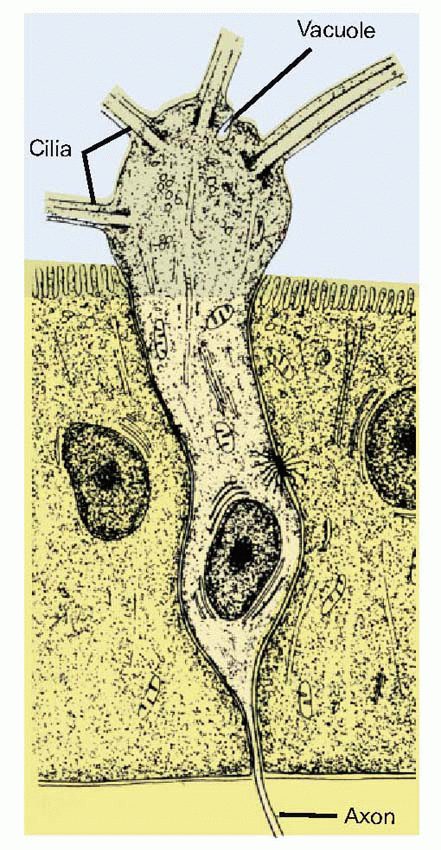
Finally there are the receptors themselves, of which there are some 10 million in Man. They are distinguished by a terminal enlargement above the surface of the epithelium, from which project some 8-20 olfactory cilia. (Note that the cilia are truncated in the figure overleaf: in practice they vary considerably in length, some being shorter than the receptor cell body, and some several times its length.) These cilia show the usual 9+2 fibril arrangement at the base, but are not believed to be actively motile: they form a dense and tangled mat that covers the olfactory area. Vacuoles can also be seen in the terminal enlargement, and experiments with exogenous protein markers have shown that they are actively pinocytotic: fluid is being continually drunk in by the receptors and passed down the olfactory nerves into the brain. The significance of this surprising feature is unclear. A further curiosity of the olfactory receptors is their remarkably short life: after a month or two at most they degenerate and are replaced by new ones from below.
 |
In addition to the conventional olfactory epithelium, there is another region, much less studied, that appears to contribute to olfaction. This is the vomeronasal organ, or Jacobson’s organ, connected by a duct to the nasal airways and probably more receptive than the olfactory epithelium to the pheromones, which are air- or water-borne signals that can have profound effects on behaviour, but in humans it exists only in the embryo (see p. 177). 
Olfactory bulb
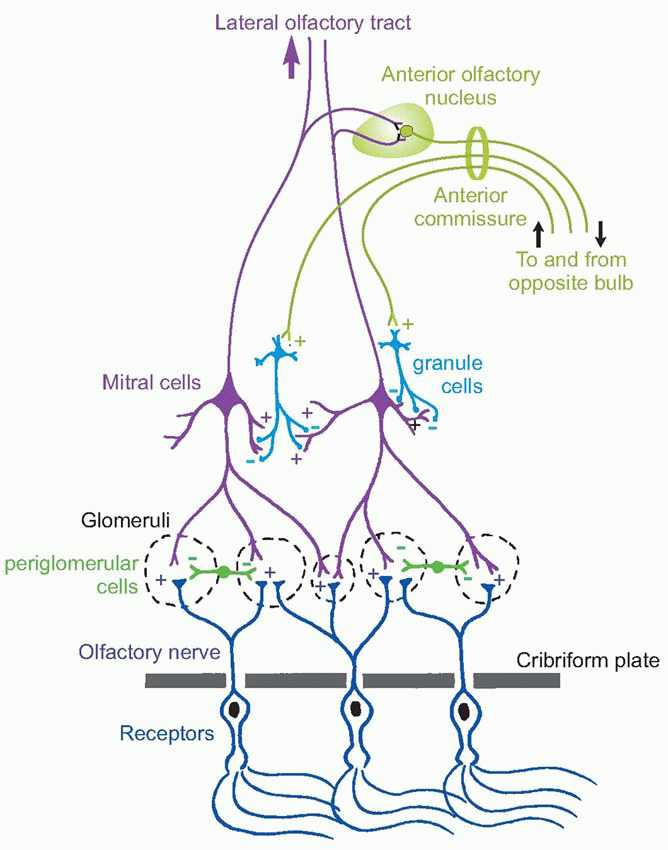
Unlike many receptor cells, the olfactory receptors send their own axons to the central nervous system (CNS) without an intervening synapse. These fibres, the fila olfactaria, make up the first cranial nerve: they are exceedingly fine and difficult to see with the light microscope: there are also vulnerable to injury from blows to the head. They pass through the cribriform plate at the base of the cranial cavity in individual holes (‘cribriform’ = ‘sieve-like’) and enter the olfactory bulb, which lies just above the olfactory epithelium. Here they synapse with dendrites of the large mitral cells (they are supposed to look like bishops’ mitres) and tufted cells in specialized nexuses called glomeruli. In rabbits, which have been particularly well studied, each glomerulus receives information from some 26 000 receptors, and has an output to about 24 mitral cells: there are probably only some 2000 glomeruli in all. The fibres entering any one glomerulus come from a wide area of the epithelium, so that detailed information about any spatial patterns of activity must be largely thrown away. But as is the case for rods in the eye, this enormous degree of convergence must certainly enhance the nose’s sensitivity by providing a mechanism by which the contributions of very large numbers of receptors can be added together. The final output of the bulb consists of the axons of the mitral and tufted cells, forming the olfactory tract: this has two divisions, lateral and medial, of which only the lateral one appears to be important in Man. In those species where it exists, the vomeronasal organ has its own nerve to an outlier of the olfactory bulb, the accessory olfactory bulb, whose projections are broadly similar to those of the main bulb itself, except that it has more prominent projections that go indirectly to the hypothalamus, as might be expected from its role in responding to pheromones.
 |
Clinical box 8.1 Anosmia after head injury
Following severe traumatic brain injury, up to 30 per cent of survivors will have anosmia. The underlying mechanisms are multiple, but shearing of the primary olfactory neurons, as they pass through the cribriform plate on their way from the nasal cavity into the cranial vault, is probably the most common cause. The sense of smell may recover, usually at 6-18 months; in some cases the anosmia is permanent, perhaps as a result of the formation of scar tissue at the cribriform plate, which prevents regenerating primary axons from reaching the secondary neurons of the olfactory bulb.
The olfactory bulb is not just a simple relay. It has two other properties that we have already seen to be common to all the sensory systems examined so far, namely lateral inhibition, and control of afferent information through negative feedback. The most prominent feedback path is formed by projections from the second-order mitral and tufted cells, which excite granule cells and which in turn inhibit neighbouring second-order cells. An interesting feature of these synapses is that they are dendro-dendritic (the granule cells have no axons), and reciprocal: excitation and inhibition occur simultaneously – in opposite directions – at the same synapse. Other collaterals of the second-order cells’ axons synapse in the anterior olfactory nucleus with interneurons that also return and synapse with granule cells; but in this case some of the interneurons project contralaterally in the anterior commissure to influence the opposite bulb in the same way. The significance of this mutual inhibition between the two bulbs is not clear: one possibility is that it may serve to enhance differences between the activities of the two bulbs – another kind of lateral inhibition – in a way that might perhaps be useful for localizing smells. Careful experiments have shown that even humans are capable of localizing odorous objects in a rather approximate way, presumably through slight differences in the timing or intensity of the stimuli in each nostril.  Finally there are the periglomerular cells, which appear to carry out local lateral inhibition at the level of the glomeruli, through reciprocal synapses with the second-order cells, and direct connections from the fila olfactaria. All this is strikingly reminiscent of the retina, where the horizontal cells appear to carry out similar functions to periglomerular cells, and amacrine cells to granule cells, using the similar kinds of synaptic mechanism. A similar arrangement is seen in the early stages of both somatosensory and auditory processing, suggesting a basic mechanism carrying out a fundamentally important function.
Finally there are the periglomerular cells, which appear to carry out local lateral inhibition at the level of the glomeruli, through reciprocal synapses with the second-order cells, and direct connections from the fila olfactaria. All this is strikingly reminiscent of the retina, where the horizontal cells appear to carry out similar functions to periglomerular cells, and amacrine cells to granule cells, using the similar kinds of synaptic mechanism. A similar arrangement is seen in the early stages of both somatosensory and auditory processing, suggesting a basic mechanism carrying out a fundamentally important function.
Finally there are the periglomerular cells, which appear to carry out local lateral inhibition at the level of the glomeruli, through reciprocal synapses with the second-order cells, and direct connections from the fila olfactaria. All this is strikingly reminiscent of the retina, where the horizontal cells appear to carry out similar functions to periglomerular cells, and amacrine cells to granule cells, using the similar kinds of synaptic mechanism. A similar arrangement is seen in the early stages of both somatosensory and auditory processing, suggesting a basic mechanism carrying out a fundamentally important function.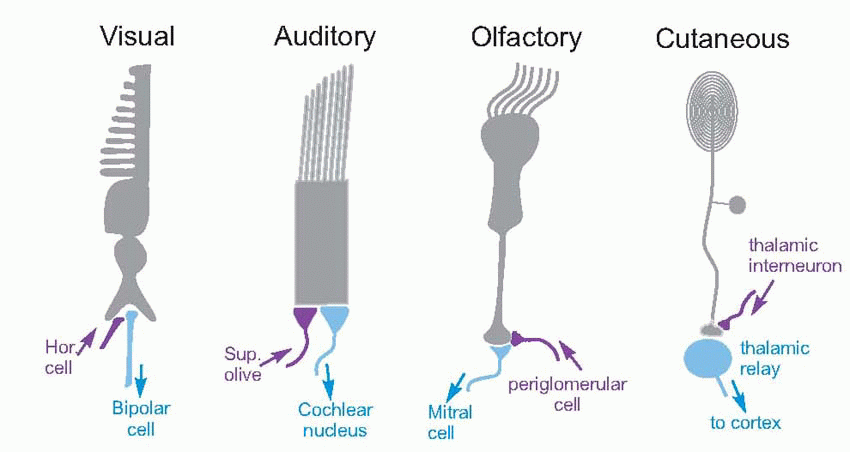 |
Central olfactory projections
The central projections of the olfactory system provide something of an embarras de richesses, very different, for example, from the visual system, with its orderly projection of the optic tract on to the lateral geniculate nucleus and thence to the cerebral cortex. Indeed olfaction seems to be unique in projecting straight on to cortical areas without relaying in the thalamus or any equivalent structure. You need to bear in mind that the olfactory system is very much older than such senses as vision and hearing, and in the course of evolution the relative size of the olfactory bulb (shown below in blue) has declined. Correspondingly, in more primitive animals a very much larger proportion of the brain is directly or indirectly concerned with olfaction. The reason for this is not hard to see: simple animals depend much more immediately than we do on knowing directly from their senses whether food is in the vicinity, and their motor systems are likely to be more pressingly governed by the need to move towards nutrients and avoid poisons, and to seek out mates by recognizing the chemical attractants they release. Their motivation – the drive that tells their motor systems what to do – is essentially olfactory.
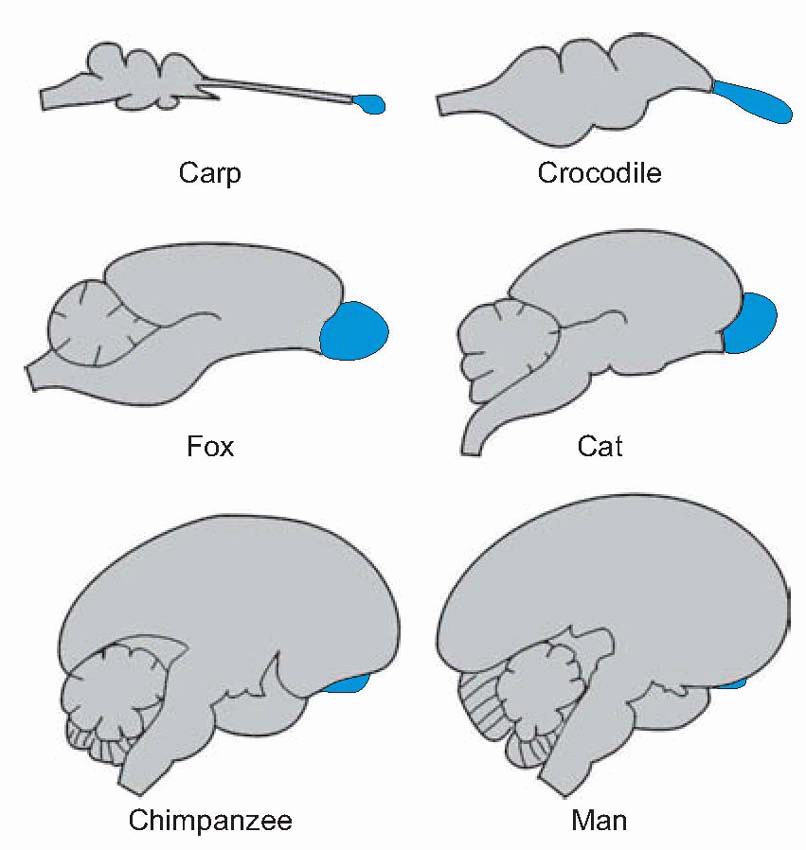 |
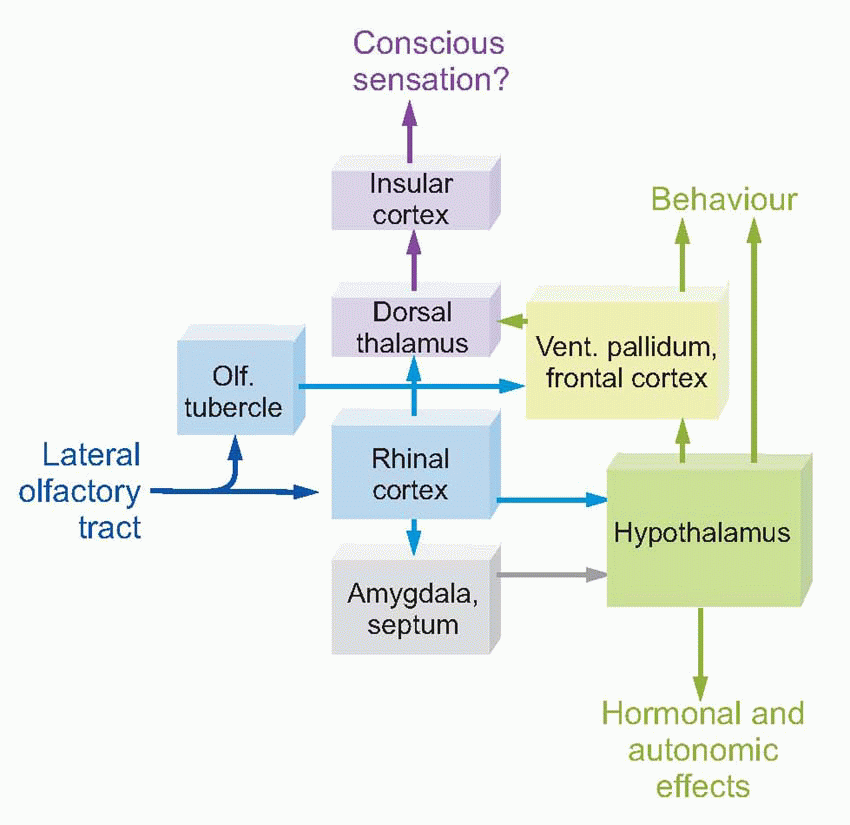 |
Even in humans, the remains of this very basic system for chemical motivation and emotion (emotion being the sensory correlate of motivation) can still be seen in the central olfactory projections. Many of these structures form part of the limbic system, a group of nuclei, cortical regions and connecting tracts of great evolutionary antiquity that appear to be concerned with precisely those kinds of function that one would expect in a primitive animal to be closely related to chemical stimulation: motivation, emotion and certain kinds of memory. The limbic system and its functions are discussed more fully in Chapters 13 and 14; for the moment we may note, for example, that the septal nuclei and amygdala contain regions known as ‘pleasure centres’, in the sense that when electrically stimulated they may provide a kind of direct positive motivation. ‘Rhinal cortex’ here includes periamygdaloid, prepyriform and entorhinal cortex, hippocampus and subiculum, areas whose functions are partly olfactory, but mostly to do with associational memory. The hippocampus in particular seems to be concerned with associating different places in the environment with the promise of food or pleasure signalled more directly by olfactory stimulation, and recognizing such a stimulus in the future as a source of motivation in its own right (see Chapter 14, p. 276).
What seems to have happened in the course of evolution is that this kind of secondary motivation by stimuli that only acquire their meaning through experience and learning has steadily grown in importance relative to that of primary olfactory motivation. For Man, money is perhaps the most obvious secondary motivator, and the most powerful of all: given the choice between a plate of fish and chips and a plate of £10 notes, unless one was exceptionally hungry there is no doubt which would cause the greater motivational drive.  For this reason, limbic structures that were originally subservient to olfaction are now not primarily olfactory at all, so the name rhinencephalon (‘nose-brain’) which is sometimes given to the limbic system is inappropriate in higher animals.
For this reason, limbic structures that were originally subservient to olfaction are now not primarily olfactory at all, so the name rhinencephalon (‘nose-brain’) which is sometimes given to the limbic system is inappropriate in higher animals.
For this reason, limbic structures that were originally subservient to olfaction are now not primarily olfactory at all, so the name rhinencephalon (‘nose-brain’) which is sometimes given to the limbic system is inappropriate in higher animals.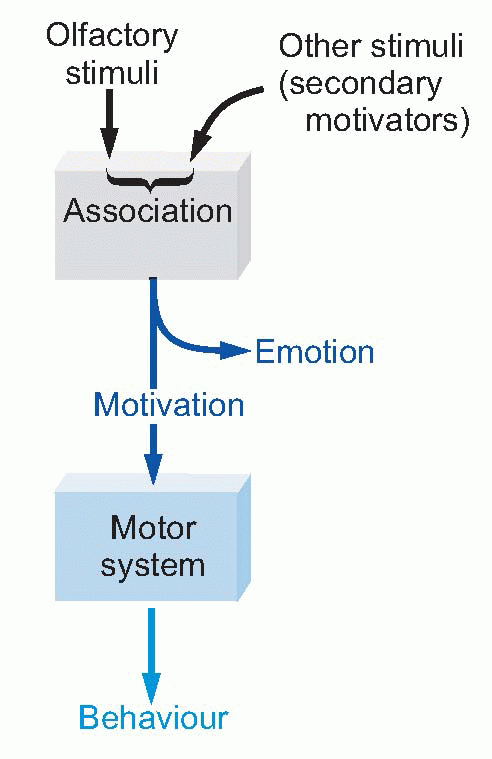 |
Finally, there are important connections between the limbic system and the hypothalamus (see Chapter 14, p. 276), providing routes by which olfactory stimuli can cause such obvious autonomic effects as salivation and other secretory responses to food smells, as well as influencing the choice of food: a single instance of particular odorant, associated with nausea even some hours later, can condition an animal to avoid the substance for the rest of its life.  Smell may also have much more widespread hormonal and behavioural effects, of which the best known are perhaps sexual arousal and modification of reproductive cycles, even abortion. A curious feature of the pheromones deployed by many creatures as long-distance sexual attractants is that they often resemble, in species as different as the civet and the moth, the steroid reproductive hormones themselves (at least in overall shape, as can be seen above). Furthermore, some of them – such as civet oil and musk – are used in perfumery and so presumably act as lures for human males as well (and they are curiously abundant also in church incense). There are said to be marked sex differences in the olfactory thresholds for some of these macrocyclic compounds, which may also vary with the phase of the menstrual cycle. But it is not just sex-life that is dominated by smell. Predators sniff out their prey; fish find their way back to their native streams – sometimes from hundreds of miles away – through smell. Pheromones in the urine or from facial glands may be used as territorial markers; herd animals use them to warn others of the approach of a predator and personal smells are an essential means by which many animals recognize their own social groups and mothers their young.
Smell may also have much more widespread hormonal and behavioural effects, of which the best known are perhaps sexual arousal and modification of reproductive cycles, even abortion. A curious feature of the pheromones deployed by many creatures as long-distance sexual attractants is that they often resemble, in species as different as the civet and the moth, the steroid reproductive hormones themselves (at least in overall shape, as can be seen above). Furthermore, some of them – such as civet oil and musk – are used in perfumery and so presumably act as lures for human males as well (and they are curiously abundant also in church incense). There are said to be marked sex differences in the olfactory thresholds for some of these macrocyclic compounds, which may also vary with the phase of the menstrual cycle. But it is not just sex-life that is dominated by smell. Predators sniff out their prey; fish find their way back to their native streams – sometimes from hundreds of miles away – through smell. Pheromones in the urine or from facial glands may be used as territorial markers; herd animals use them to warn others of the approach of a predator and personal smells are an essential means by which many animals recognize their own social groups and mothers their young. 
Smell may also have much more widespread hormonal and behavioural effects, of which the best known are perhaps sexual arousal and modification of reproductive cycles, even abortion. A curious feature of the pheromones deployed by many creatures as long-distance sexual attractants is that they often resemble, in species as different as the civet and the moth, the steroid reproductive hormones themselves (at least in overall shape, as can be seen above). Furthermore, some of them – such as civet oil and musk – are used in perfumery and so presumably act as lures for human males as well (and they are curiously abundant also in church incense). There are said to be marked sex differences in the olfactory thresholds for some of these macrocyclic compounds, which may also vary with the phase of the menstrual cycle. But it is not just sex-life that is dominated by smell. Predators sniff out their prey; fish find their way back to their native streams – sometimes from hundreds of miles away – through smell. Pheromones in the urine or from facial glands may be used as territorial markers; herd animals use them to warn others of the approach of a predator and personal smells are an essential means by which many animals recognize their own social groups and mothers their young. 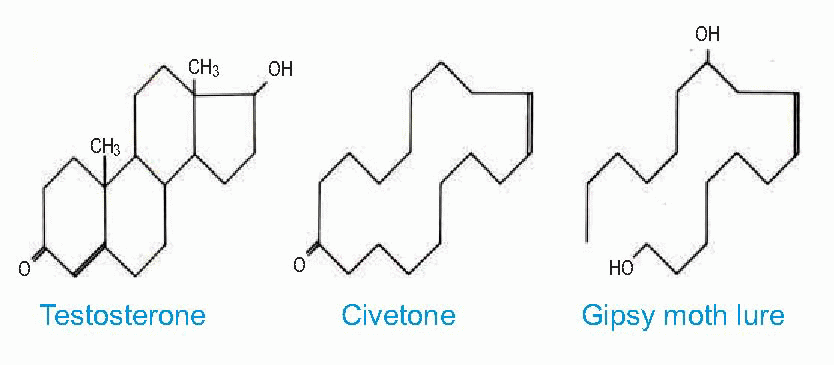 |
It is natural to speculate whether we humans are influenced subconsciously in the same kind of way: if so, it would have the most profound psychological and sociological implications. Firm evidence is lacking, though pheromone sprays for men apparently sell well.  The vomeronasal organ, when it exists, appears to be particularly involved in detecting olfactory stimuli with strong behavioural implications: via the accessory olfactory bulb, it projects extensively to various parts of the limbic system, particularly the medial nucleus of the amygdala (Table 8.1).
The vomeronasal organ, when it exists, appears to be particularly involved in detecting olfactory stimuli with strong behavioural implications: via the accessory olfactory bulb, it projects extensively to various parts of the limbic system, particularly the medial nucleus of the amygdala (Table 8.1).
The vomeronasal organ, when it exists, appears to be particularly involved in detecting olfactory stimuli with strong behavioural implications: via the accessory olfactory bulb, it projects extensively to various parts of the limbic system, particularly the medial nucleus of the amygdala (Table 8.1).One striking indication of the psychological links between olfactory and limbic functions in Man is the very vivid way in which odours may call up – often with surprising intensity – recollections of past experience; and it is significant that so often such evocations are not just of
the objective circumstances of a particular event, but also of the mood or emotion that was felt at the time, in a way that is seldom experienced with purely auditory or visual stimulation. The direct penetration of the emotional areas of the brain by olfactory fibres seems exactly reflected in what we feel.
the objective circumstances of a particular event, but also of the mood or emotion that was felt at the time, in a way that is seldom experienced with purely auditory or visual stimulation. The direct penetration of the emotional areas of the brain by olfactory fibres seems exactly reflected in what we feel.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


