Figure 26-1. A series of stacked boxes (A) illustrating which nuclei form the various parts of the basal nuclei and how these groups are used in this chapter (B). A standard drawing of the basal nuclei (C) used throughout this chapter.
The telencephalic regions of the basal nuclei are supplied by the medial striate artery, the lenticulostriate branches of the M1 segment of the middle cerebral artery, and the anterior choroidal artery (Fig. 26-2). The diencephalic and mesencephalic regions are supplied by the posteromedial branches of the P1 segment of the posterior cerebral artery and branches of the posterior communicating artery. Diseases of these vessels may result in various behavioral or motor deficits, depending on which vessel and region are affected.

Figure 26-2. Blood supply to the basal nuclei in coronal (A) and axial (B) planes. The anterior choroidal artery serves the more inferior aspects of the genu and posterior limb; the more superior aspects are served by the lenticulostriate arteries.
Striatal Complex
The striatal complex is a functional unit composed of the neostriatum and ventral striatum (Fig. 26-3A, B). The neostriatum consists of the caudate nucleus and putamen. These two nuclei have the same embryologic origin and similar connections. Although fused rostroventrally, they are separated throughout most of their extents by fibers of the internal capsule. The ventral striatum is composed of the nucleus accumbens and portions of the olfactory tubercle (Figs. 26-1A, B and 26-3A). The nucleus accumbens is located rostroventrally in the hemisphere, at the point where the putamen is continuous with the head of the caudate (Figs. 26-3A and 26-4A, B). It is internal to part of the anterior perforated substance. Portions of the olfactory tubercle are considered part of the ventral striatum because of functional, cytoarchitectural, and chemoarchitectural similarities. A defining characteristic of the striatal complex, patches (also called striosomes) are particularly prominent in the head of the caudate (Fig. 26-4B). Patches are acetylcholinesterase-poor regions within the striatal complex. They contain large amounts of one or more neuropeptides and one or more types of opiate receptors. Patches are surrounded by matrix (Fig. 26-4B), which contains high concentrations of acetylcholine and therefore stains darkly when tissue is histochemically reacted to reveal this enzyme. In addition to histochemical and receptor differences, these regions also receive projections from different cortical regions and project to different targets.

Figure 26-3. Cross sections of the human brain from rostral (A) to caudal (D) showing the basal nuclei and related structures. Myelin stain. (A Modified from Haines DE: Neuroanatomy: An Atlas of Structures, Sections, and Systems, 8th ed. Baltimore, Lippincott Williams & Wilkins, 2012.)

Figure 26-4. Cross section of the rostral and basal forebrain showing the location of the nucleus accumbens at the continuation of the head of the caudate nucleus with the putamen (A). The main afferent projections to and efferent projections from the nucleus accumbens are represented in B.
The largest afferent projections to the neostriatum are from the cerebral cortex (corticostriatal fibers) (Fig. 26-5A). Other afferents are from the thalamus (thalamostriatal fibers), substantia nigra (nigrostriatal fibers), and parabrachial pontine reticular formation (pedunculopontostriatal fibers) (Fig. 26-5A). The efferent projections of the striatum reach primarily the pallidum (striatopallidal fibers) and the nigral complex (striatonigral fibers) and to a small degree the subthalamic nucleus.
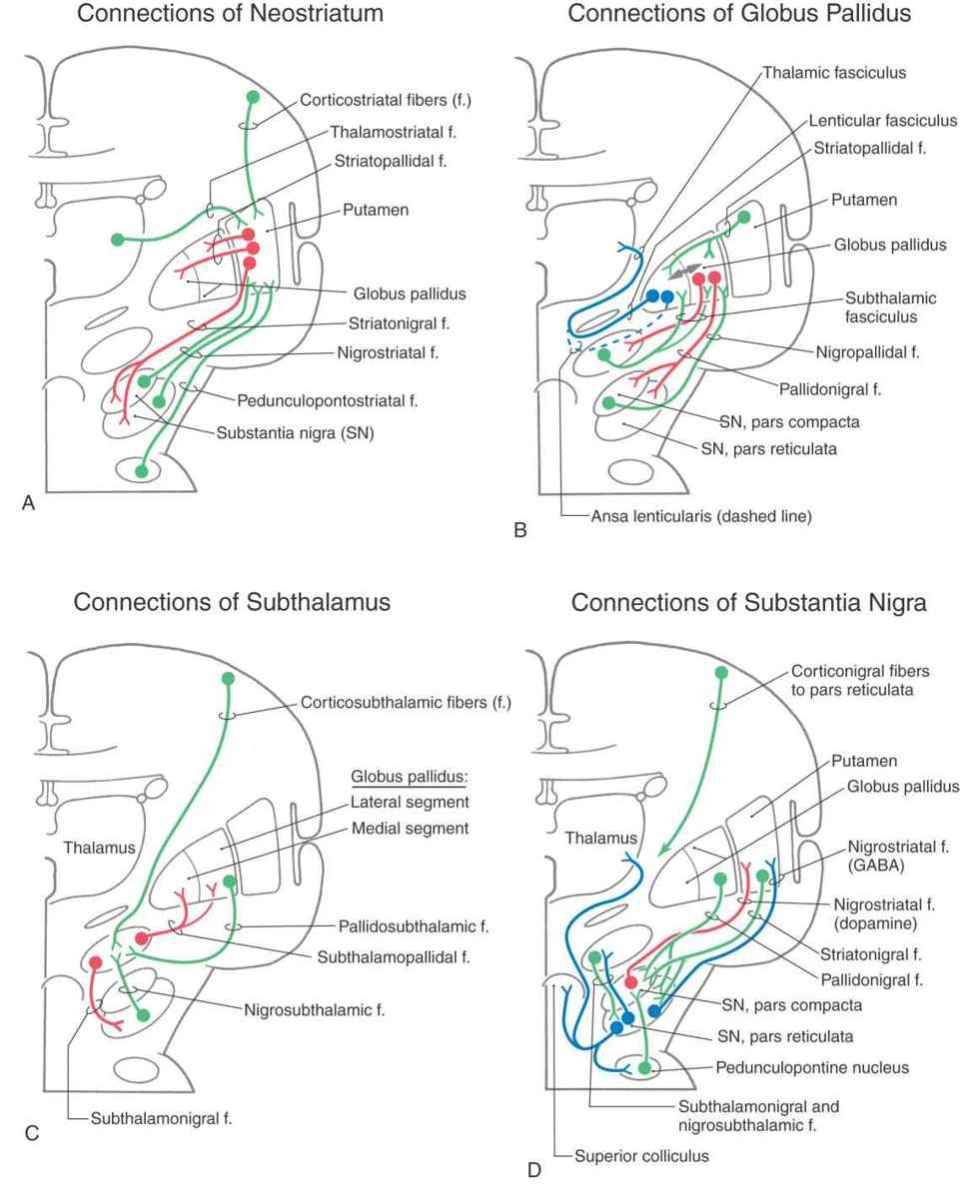
Figure 26-5. Schematic representations of the afferent (in green) and efferent (in red) connections of the neostriatum (A) and subthalamus (C) and of the afferent (in green) and efferent (in red and blue) connections of the globus pallidus (B) and substantia nigra (D). The double-headed arrow in B represents pallidopallidal fibers.
Most of the neurons in the neostriatum are called medium spiny neurons, so named because of their medium-sized cell bodies and the large numbers of spines on their dendrites (Fig. 26-6). Most medium spiny cells have dendritic fields that are restricted to the patch or matrix compartment in which the cell bodies are located.

Figure 26-6. Medium spiny neuron from the primate neostriatum. The detail shows the characteristic appearance of dendritic spines on these cells. (Photos courtesy of Dr. José Rafols.)
The nucleus accumbens forms the majority of the ventral striatum (Fig. 26-4A). This nucleus is divided into a core region and a shell region. The core region is cytoarchitecturally and histochemically identical to the neostriatum. It has similar efferent and afferent connections, albeit from cortical regions and to different pallidal and midbrain nuclei. The shell is somewhat histochemically different and has a more diffuse set of connections. It receives projections primarily from allocortical regions and sends projections to the ventral pallidum, substantia nigra–ventral tegmental area, parabrachial nucleus, periaqueductal gray, lateral hypothalamus, and lateral preoptic area (Fig. 26-4B).
Medium spiny neurons fire few action potentials spontaneously and thus require activation by their afferent fibers. These cells use the inhibitory neurotransmitter γ-aminobutyric acid (GABA) and may also contain neuroactive peptides such as substance P and enkephalin. Thus when medium spiny neurons are activated, they subserve both direct inhibitory and neuromodulatory functions at their targets. The axons of medium spiny neurons are the efferent fibers of the neostriatum, collectively forming the striatopallidal fibers.
Also found in the neostriatum are large, acetylcholine-containing local circuit neurons that modulate local activity within the neostriatum. Huntington disease is characterized by progressive loss of medium spiny neurons and acetylcholine-containing neurons throughout the striatal complex.
Pallidal Complex
The pallidal complex is composed of the globus pallidus and the ventral pallidum. The latter is largely synonymous with the substantia innominata (Fig. 26-3B, C). The pallidal complex contains primarily GABAergic neurons with high rates of spontaneous activity. Consequently, these cells tonically inhibit their targets.
The globus pallidus is divided into medial (internal) and lateral (external) segments by a sheet of white matter (the medullary lamina) (Fig. 26-3C). The substantia innominata is located inferior to the anterior commissure and internal to the anterior perforated substance. One important cell group in the substantia innominata is the basal nucleus of Meynert. This nucleus has large acetylcholine-containing neurons, which are lost in Alzheimer disease. However, this disease is not considered a basal nuclear disorder because acetylcholine-containing cells in the cerebral cortex, hippocampus, and septum are also lost in patients with Alzheimer disease. This disease is further characterized by other biochemical and pathologic features, such as senile plaques.
The two divisions of the globus pallidus are reciprocally connected (pallidopallidal fibers) (Fig. 26-5B) but subserve different functions. The main afferent input to the pallidum is from the striatal complex. Medium spiny neurons from the striatum that project to the medial segment and substantia nigra use GABA and substance P; those that project to the lateral segment use GABA and enkephalin (Fig. 26-5B).
The medial division is composed of the medial segment of the globus pallidus. It subserves the direct basal nuclear pathway (described later) and projects primarily to the thalamus (pallidothalamic fibers) (Fig. 26-5B). These fibers exit the globus pallidus as two bundles: the ansa lenticularis and the lenticular fasciculus (Figs. 26-3C, D; see also Figs. 26-8 and 26-9). The ansa lenticularis originates from lateral portions of the medial segment and loops around the posterior limb of the internal capsule to enter the prerubral field (field H of Forel). The lenticular fasciculus (field H2 of Forel), on the other hand, originates in the posteromedial portion of the medial segment. These fibers traverse the internal capsule as small groups of axons, merge to form the lenticular fasciculus between the zona incerta and subthalamic nucleus, and then enter field H of Forel. In the Forel field, the ansa lenticularis and lenticular fasciculus join the thalamic fasciculus (field H1 of Forel), which courses immediately superior to the zona incerta (Fig. 26-3C, D; see also Figs. 26-8 and 26-9). These fibers ultimately terminate in ventral anterior, ventral lateral, and centromedian nuclei of the thalamus. The medial division of the pallidal complex is a principal efferent nucleus of the basal nuclei, the axons of these cells comprising the ansa lenticularis and the lenticular fasciculus.
The lateral division is composed of the external (or lateral) segment of the globus pallidus and the ventral pallidum. This division subserves the indirect basal nuclear pathway (see later). These nuclei receive a large input from the striatal complex (striatopallidal fibers) and small projections from the subthalamic nucleus (subthalamopallidal fibers) and the substantia nigra pars reticulata (nigropallidal fibers). They project strongly to the subthalamic nucleus (pallidosubthalamic fibers) and are also connected with the substantia nigra (pallidonigral fibers) (Fig. 26-5B).
Subthalamic Nucleus
The subthalamic nucleus is a lens-shaped cell group that makes up the largest part of the ventral thalamus. It is immediately inferior to the zona incerta and rostral to the substantia nigra (Fig. 26-3C, D). It receives projections from the lateral pallidal division (pallidosubthalamic fibers), cerebral cortex (corticosubthalamic fibers), nigral complex (nigrosubthalamic fibers), and parabrachial pontine reticular formation. The subthalamic nucleus projects to both pallidal divisions (subthalamopallidal fibers) and to the substantia nigra (subthalamonigral fibers) (Fig. 26-5C). These connections, especially the subthalamopallidal projections to the medial globus pallidus, are an essential part of the indirect pathway underlying basal nuclear function.
Subthalamic neurons use the excitatory neurotransmitter glutamate. Most of the time, subthalamic cells are inactive because of the constant inhibition by cells of the external pallidal segment. However, if this inhibition is removed, subthalamic neurons have a high level of activity resulting in a characteristic motor deficit described later in this chapter. This activity is mediated in part by a large corticosubthalamic projection.
Nigral Complex
The nigral complex is composed of the substantia nigra and the ventral (anterior) tegmental area (Fig. 26-3C, D). The substantia nigra is divided into a cell-dense portion (pars compacta) and a reticulated portion (referred to here as the pars reticulata, although it can be divided into a pars reticulata and a pars lateralis). The pars reticulata is located at and within the medial edge of the descending corticofugal fibers that form the crus cerebri. The pars compacta and the adjacent ventral tegmental area appear to subserve similar functions and to have a similar chemoarchitectural organization. The major afferents to the nigral complex are from the striatal and pallidal complexes. The nigral complex also receives cortical (corticonigral), subthalamic (subthalamonigral), and pedunculopontine fibers (Fig. 26-5D).
The pars compacta contains a large number of neuromelanin-containing cells, whose dark color gives the nucleus its name (substantia nigra, “black substance”). Cells of the pars compacta in healthy individuals appear characteristically dark in brain slices and are packed with small black granules in histologic sections (Fig. 26-7A–C). Neurons in the pars compacta use the neurotransmitter dopamine and project primarily to the neostriatum as nigrostriatal fibers. The dopamine released by these cells may excite or inhibit striatal neurons, depending on the type of receptor on the postsynaptic membrane.

Figure 26-7. Axial slice of the midbrain showing the substantia nigra in a normal brain (A) in its characteristic position immediately internal to the crus cerebri. Histologic sections at low-power (B) and high-power (C) magnification reveal the dense packing of melanin in normal neurons. In a patient with Parkinson disease, nigral neurons have little or no melanin (D, arrows), with consequent loss of dopamine, and Lewy bodies of various sizes are found on the neuronal cytoplasm (D). (Specimens in C and D courtesy of Dr. Jonathan Fratkin.)
The pars reticulata is formed by loose aggregations of medium-sized to large GABAergic neurons that are indistinguishable from those of the medial pallidum. Neurons in the pars reticulata have axons with an extensive system of collaterals; consequently, they may project to and inhibit one or more target structures. These targets include the neostriatum (nigrostriatal fibers), thalamus (nigrothalamic fibers), superior colliculus (nigrotectal fibers), and parabrachial pontine reticular formation. These cells have a high rate of discharge and tonically inhibit their targets. Projections of pars reticulata neurons represent an important pathway by which the basal nuclei influence other motor centers.
The pars compacta and the pars reticulata are interconnected. Dendrites of dopaminergic pars compacta neurons extend into the pars reticulata, where they release free dopamine (by a nonvesicular mechanism). The level of dopamine modulates the resting membrane potential of pars reticulata cells, making them either more or less likely to discharge, depending on the subtype of dopamine receptor they possess. In turn, pars reticulata neurons have axon collaterals that ramify extensively in the pars compacta and form GABAergic synapses. Collectively, these interactions form modulatory loops between neurons of the pars compacta and the pars reticulata.
These modulatory loops are influenced by the output of the striatal system. Neurons in the patches project predominantly onto the cells in the pars compacta, whereas the neurons in the matrix project predominantly onto the cells in the pars reticulata. Thus the striatonigral projection from patches directly inhibits the dopaminergic nigrostriatal neurons, and the projection from the matrix inhibits the GABAergic neurons in the pars reticulata. Consequently, inhibition of the pars reticulata is reduced and its targets, such as the pars compacta, are released from inhibition and may become more active.
The ventral tegmental area is located medial to the substantia nigra. It contains large numbers of dopaminergic neurons and forms connections with the ventral striatum, the amygdala, and other limbic system structures. Cells of the ventral tegmental area project to and terminate on striatal neurons that have postsynaptic D2 (dopamine) receptors. In schizophrenia, there is an increase in number and in sensitivity of these receptors. Neuroleptic drugs help control schizophrenia by blocking (downregulating) these D2 receptors.
Parabrachial Pontine Reticular Formation
Nuclei in the region of the parabrachial pontine reticular formation, primarily the pedunculopontine tegmental nucleus, are intimately connected with all portions of the basal nuclei and their associated nuclei. For example, GABAergic substantia nigra pars reticulata neurons project onto cells of the pedunculopontine tegmental nucleus. In turn, acetylcholine-containing pedunculopontine tegmental neurons project to the substantia nigra pars compacta. In addition, the pedunculopontine tegmental nucleus is reciprocally connected with the subthalamic nucleus. Moreover, the parabrachial pontine nuclei are connected with motor centers in the brainstem, which project to the spinal cord via the descending spinal pathways. Thus these nuclei serve as an efferent pathway for the basal nuclei. It has been suggested that damage to the connections between the pedunculopontine nucleus and the basal nuclei may partially account for motor deficits, such as tremor or chorea, in some patients.
Ventral Basal Nuclei
The ventral basal nuclear pathways are similar to those of the dorsal basal nuclei. The pathways originate in the allocortex (limbic-related cortical areas) and the orbital and medial prefrontal cortices. The corticostriatal pathway terminates in the ventral striatum (nucleus accumbens and portions of the olfactory tubercle). Striatopallidal neurons then project to the ventral pallidum (substantia innominata). The pallidothalamic neurons then project to the mediodorsal nucleus of the thalamus, whose neurons then project to the cerebral cortex. As with the dorsal basal nuclei, distinct regions of each nucleus in these pathways are connected with each other. Thus there is a clear loop through the ventral basal nuclei for information originating from different areas of the cortex, as is discussed later.
DIRECT AND INDIRECT PATHWAYS OF BASAL NUCLEAR ACTIVITY
Pathways through the basal nuclei consist of parallel circuits that share certain features. The basic circuit is divided into direct and indirect pathways that have opposing actions on targets of the basal nuclei (Figs. 26-8 to 26-10). As a general concept, the direct pathway facilitates a flow of information through the thalamus and the indirect pathway inhibits this flow. These pathways create a balance in the inhibitory outflow of the basal nuclei and function by modulating the extent of this inhibition on target nuclei.

Figure 26-8. Schematic representation of pallidal connections to the thalamus and with the subthalamic nucleus.

Figure 26-9. Three-dimensional representation of the connections of the basal nuclei, excluding those with the substantia nigra. Fibers of the thalamic fasciculus (dark blue) traverse the internal capsule, pass between the subthalamic nucleus and zona incerta, and then enter the thalamic fasciculus to access the VLpo. Fibers of the ansa lenticularis (light blue) arch around the rostromedial edge of the internal capsule and course caudally to enter the thalamic fasciculus en route to the VLpo. Pallidosubthalamic and subthalamopallidal fibers (both green) collectively form the subthalamic fasciculus. Cerebellothalamic fibers from the contralateral cerebellar nuclei also use the thalamic fasciculus to get to the VLpc. VLpc, ventral lateral thalamic nucleus, pars caudalis; VLpo, ventral lateral thalamic nucleus, pars oralis. (Modified from Nieuwenhuys R, Voogd J, van Huijzen C: The Human Central Nervous System, A Synopsis and Atlas, 3rd ed. Berlin, Springer-Verlag, 1988, with permission.)

Figure 26-10. Schematic representation of direct and indirect pathways through the basal nuclei.
The direct pathway (Figs. 26-9, 26-10, and 26-11A) begins as an excitatory, glutamatergic projection from the cerebral cortex to the striatal complex. Striatal neurons inhibit cells in the internal (or medial) segment of the globus pallidus and in the substantia nigra pars reticulata. These striatopallidal and striatonigral fibers use GABA and substance P. Cells of the internal segment of the globus pallidus (as pallidothalamic fibers
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


