SECTION IV ANATOMY OF THE BRAIN
CHAPTER 7
The Brain Stem and Cerebellum
The brain stem includes the medulla and pons, located ventral to the cerebellum. In addition to housing essential ascending and descending tracts, the brain stem contains nuclei that are essential for maintenance of life. As a result of the relatively tight packaging of numerous ascending and descending tracts, as well as nuclei, within the brain stem, even small lesions within it can injure multiple tracts and nuclei within it and thus can produce very significant neurologic deficits. The cerebellum, located just dorsal to the brain stem, plays a major role in motor coordination. Because of its proximity to the brain stem, injuries which cause swelling of the cerebellum can compress the brain stem, and thus can rapidly become life-threatening.
DEVELOPMENT OF THE BRAIN STEM AND CRANIAL NERVES
The lower part of the cranial portion of the neural tube (neuraxis) gives rise to the brain stem. The brain stem is divided into the mesencephalon and rhombencephalon (Fig 7–1).
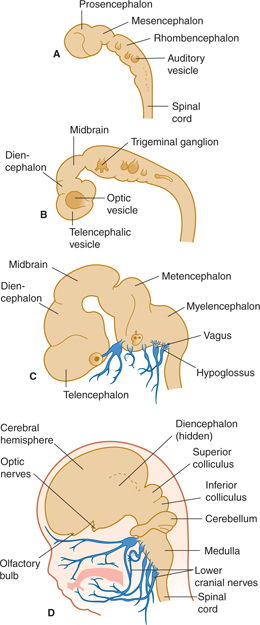
FIGURE 7–1 Four stages in early development of brain and cranial nerves (times are approximate). A: 3½ weeks. B: 4½ weeks. C: 7 weeks. D: 11 weeks.
The primitive central canal widens into a four-sided pyramid shape with a rhomboid floor (Fig 7–2). This becomes the fourth ventricle, which extends over the future pons and the medulla.
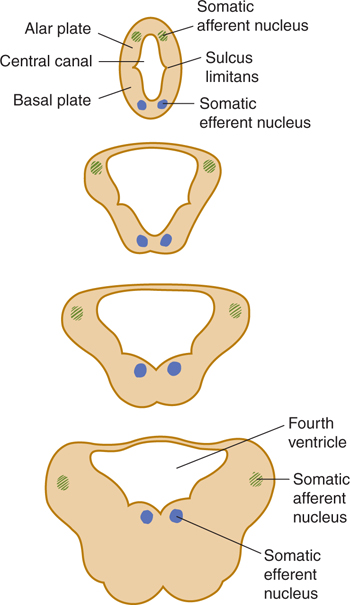
FIGURE 7.2 Schematic illustration of the widening of the central cavity in the lower brain stem during development.
The neural tube undergoes local enlargement and shows two permanent flexures: the cephalic flexure at the upper end and the cervical flexure at the lower end. The cephalic flexure in an adult brain is the angle between the brain stem and the horizontal plane of the brain (see Fig 1–6).
The central canal in the rostral brain stem becomes the cerebral aqueduct. The roof of the rostral fourth ventricle undergoes intense cellular proliferation, and this lip produces the neurons and glia that will populate both the cerebellum and the inferior olivary nucleus.
The quadrigeminal plate, the midbrain tegmentum, and the cerebral peduncles develop from the mesencephalon (midbrain; see Fig 7–1), and the cerebral aqueduct courses through it. The rhombencephalon (see Fig 7–1A) gives rise to the metencephalon and the myelencephalon. The metencephalon forms the cerebellum and pons; it contains part of the fourth ventricle. The myelencephalon forms the medulla oblongata; the lower part of the fourth ventricle lies within this portion of the brain stem.
As in the spinal cord, the embryonic brain stem has a central gray core with an alar plate (consisting mostly of sensory components) and a basal plate (composed primarily of motor components). The gray columns are not continuous in the brain stem, however, and the development of the fourth ventricle causes wide lateral displacement of the alar plate in the lower brain stem. The basal plate takes the shape of a hinge (see Fig 7–2). The process is reversed at the other end, resulting in the rhomboid shape of the floor of the fourth ventricle. In addition, long tracts, short neuronal connections, and nuclei become apposed to the brain stem. The cranial nerves, like the spinal nerves, take their origin from the basal plate cells (motor nerves) or from synapses in the alar plate cell groups (sensory nerves). Unlike spinal nerves, most cranial nerves emerge as one or more bundles of fibers from the basal or basilateral aspect of the brain stem (Figs 7–1 and 7–3). In addition, not all cranial nerves are mixed; some have only sensory components and others have only motor components (see Chapter 8).
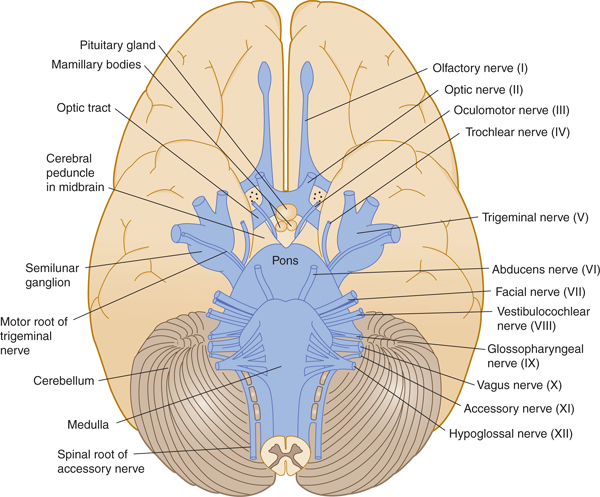
FIGURE 7–3 Ventral view of the brain stem, in relation to cerebral hemispheres and cerebellum, showing the cranial nerves.
BRAIN STEM ORGANIZATION
Main Divisions and External Landmarks
Three major external divisions of the brain stem are recognizable: the medulla (medulla oblongata), the pons together with the cerebellum, and the midbrain (mesencephalon) (Figs 7–3 and 7–4). The three internal longitudinal divisions of the brain stem are the tectum (mainly in the midbrain), tegmentum, and basis (see Fig 7–4). Thus, the pons, for example, can be considered to consist of a dorsal pontine tegmentum and a ventral basis pontis. The main external structures, seen from the dorsal aspect, are shown in FIGURE 7–5. The superior portion of the rhomboid fossa (which forms the floor of the fourth ventricle) extends over the pons, whereas the inferior portion covers the open portion of the medulla. The closed medulla forms the transition to the spinal cord.
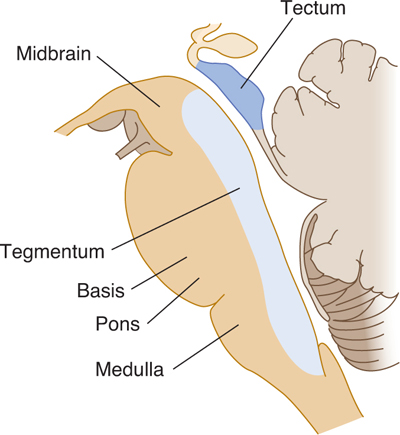
FIGURE 7–4 Drawing of the divisions of the brain stem in a midsagittal plane. The major internal longitudinal divisions are the tectum, tegmentum, and basis. The major external divisions are the midbrain, pons, and medulla.
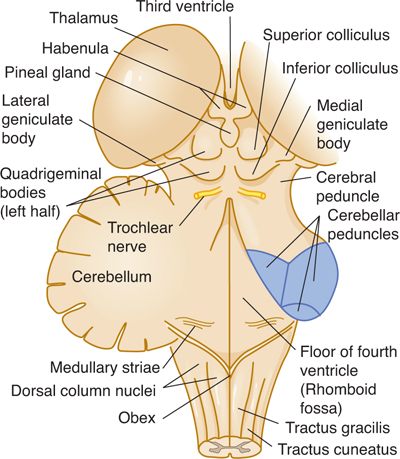
FIGURE 7–5 Dorsolateral aspect of the brain stem (most of cerebellum removed).
Three pairs of cerebellar peduncles (inferior, middle, and superior) form connections with the cerebellum. The dorsal aspect of the midbrain shows four hillocks: the two superior and the two inferior colliculi, collectively called the corpora quadrigemina or quadrigeminal plate.
Internal Structural Components
A. Descending and Ascending Tracts
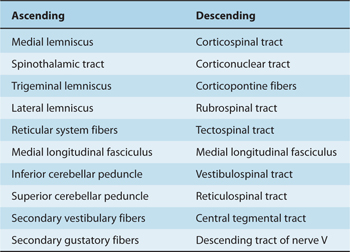
All descending tracts that terminate in the spinal cord (eg, the corticospinal tract; see Chapter 5) pass through the brain stem. In addition, several descending fiber systems terminate or originate in the brain stem. Similarly, all ascending tracts (eg, the spinothalamic tracts) that reach the brain stem or the cerebral cortex pass through part or all of this region; other ascending tracts originate in the brain stem. The brain stem is, therefore, an important conduit or relay station for many longitudinal pathways, both descending and ascending (Table 7–1).
Table 7–1 Major Ascending and Descending Pathways in the Brain Stem.
B. Cranial Nerve Nuclei
Almost all the cranial nerve nuclei are located in the brain stem. (The exceptions are the first two cranial nerve nuclei, which are evaginations of the brain itself.) Portions of the cranial nerves also pass through the brain stem.
C. Cerebellar Peduncles
The pathways to and from the cerebellum pass through three pairs of cerebellar peduncles, as described later in the Cerebellum section.
D. Descending Autonomic System Pathways
These paths to the spinal cord pass through the brain stem (see Chapter 20).
E. Reticular Formation
Several of these areas in the tegmentum of the brain stem are vitally involved in the control of respiration; cardiovascular system functions; and states of consciousness, sleep, and alertness (see Chapter 18).
F. Monoaminergic Pathways
These paths include three important systems: the serotonergic pathways from the raphe nuclei (see Chapter 3); the noradrenergic pathways in the lateral reticular formation and the extensive efferents from the locus ceruleus; and the dopaminergic pathway from the basal midbrain to the basal ganglia and others.
CRANIAL NERVE NUCLEI IN THE BRAIN STEM
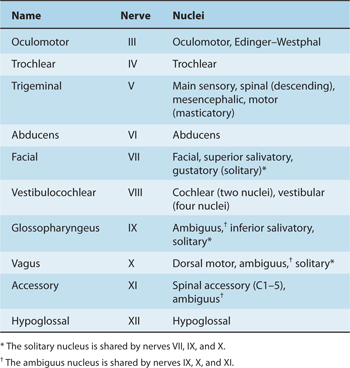
The functional composition of the lower 10 cranial nerves can be analyzed by referring to the development of their nuclei (Fig 7–6). The nerves are usually referred to by name or by Roman numeral (Table 7–2).
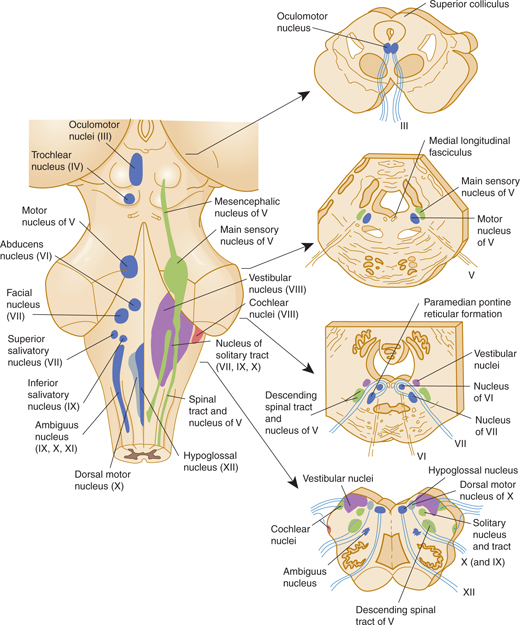
FIGURE 7–6 Cranial nerve nuclei. Left: Dorsal view of the human brain stem with the positions of the cranial nerve nuclei projected on the surface. Motor nuclei are on the left; sensory nuclei are on the right. Right: Transverse sections at the levels indicated by the arrows.
Table 7–2 Cranial Nerves and Nuclei in the Brain Stem.
Motor (Efferent) Components
Three types of basal plate derivatives (motor nuclei) are located within the brain stem (see Table 7–2).
General somatic efferent (SE or GSE) components innervate striated muscles that are derived from somites and are involved with movements of the tongue and eye, such as the hypoglossal nucleus of XII, oculomotor nucleus of III, trochlear nucleus of IV, and abducens nucleus of VI.
Branchial efferent (BE) components, sometimes referred to as special visceral efferents (SVE), innervate muscles that are derived from the branchial arches and are involved in chewing, making facial expressions, swallowing, producing vocal sounds, and turning the head. Examples include the masticatory nucleus of V; facial nucleus of VII; ambiguus nucleus of IX, X, and XI; and spinal accessory nucleus of XI located in the cord.
General visceral efferent (VE or GVE) components are parasympathetic preganglionic components that provide autonomic innervation of smooth muscles and the glands in the head, neck, and torso. Examples include the Edinger-Westphal nucleus of III, superior salivatory nucleus of VII, inferior salivatory nucleus of IX, and dorsal motor nucleus of X.
Sensory (Afferent) Components
Two types of alar-plate derivatives can be distinguished in the brain stem and are comparable to similar cell groups in the spinal cord (see Table 7–2).
General somatic afferent (SA or GSA) components receive and relay sensory stimuli from the skin and mucosa of most of the head: main sensory, descending, and mesencephalic nuclei of V.
General visceral afferent (VA or GVA) components relay sensory stimuli from the viscera and more specialized taste stimuli from the tongue and epiglottis: solitary nucleus for visceral input from IX and X and gustatory nucleus for special visceral taste fibers from VII, IX, and X.
Six special sensory (SS) nuclei can also be distinguished: the four vestibular and two cochlear nuclei that receive stimuli via vestibulocochlear nerve VIII. These nuclei are derived from the primitive auditory placode in the rhombencephalon (Fig 7–7A).
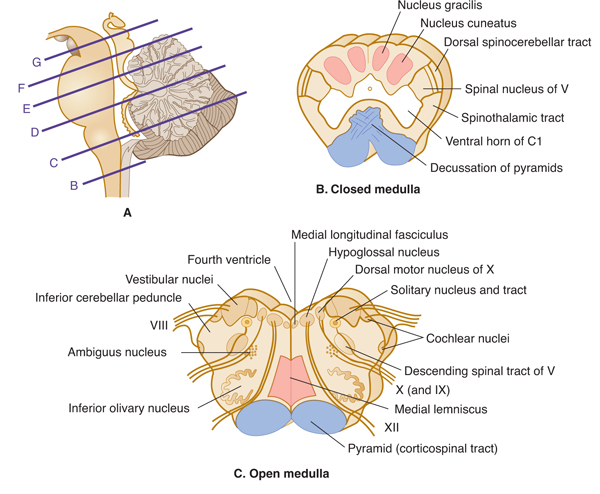
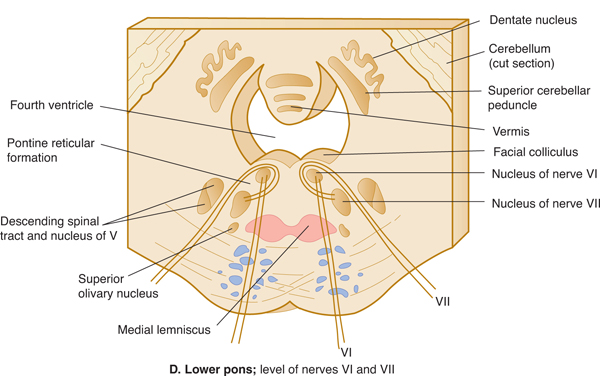
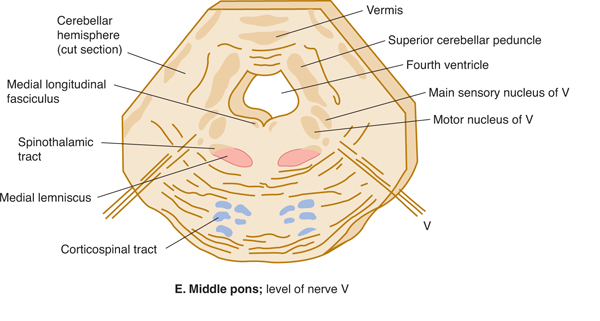
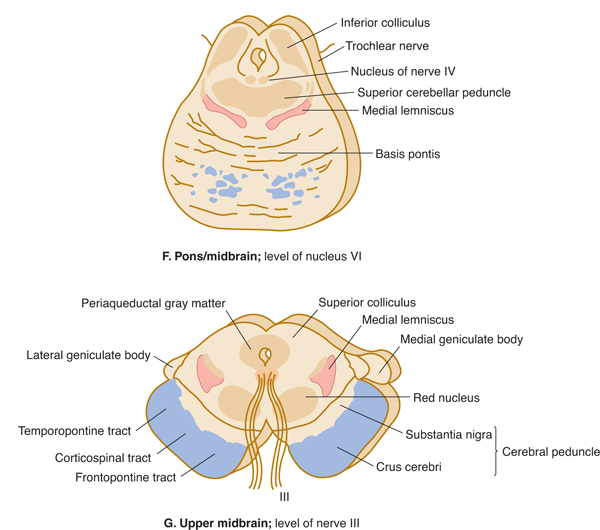
FIGURE 7–7 A: Key to levels of sections. B-G: Schematic transverse sections through the brain stem. The corticospinal tracts and the dorsal column nuclei/medial lemnisci are shown in color so that they can be followed as they course through the brain stem.
Differences Between Typical Spinal and Cranial Nerves
The simple and regular pattern of organization of spinal nerves is not found in cranial nerves. There is no single blueprint and the cranial nerves must thus be learned one-by-one. A single cranial nerve may contain one or more functional components; conversely, a single nucleus may contribute to the formation of one or more cranial nerves. Although some cranial nerves are solely efferent, most are mixed, and some contain many visceral components. The cranial nerves are described in detail in Chapter 8.
MEDULLA
The medulla (medulla oblongata) can be divided into a caudal (closed; Fig 7–7B) portion and a rostral (open; Fig 7–7C) portion. The division is based on the absence or presence of the lower fourth ventricle.
Ascending Tracts
In the caudal, closed part of the medulla, the relay nuclei of the dorsal column pathway (nucleus gracilis and nucleus cuneatus) give rise to a crossed fiber bundle, the medial lemniscus. The lower part of the body is represented in the ventral portion of the lemniscus and the upper part of the body in the dorsal. The spinothalamic tract (which crossed at spinal cord levels) continues upward throughout the medulla, as do the spinoreticular tract and the ventral spinocerebellar pathway. The dorsal spinocerebellar tract and the cuneocerebellar tract continue into the inferior cerebellar peduncle.
Descending Tracts
The corticospinal tract in the pyramid begins to cross at the transition between medulla and spinal cord; this decussation takes place over several millimeters. Most of the axons in this tract arise in the motor cortex. Some fibers from the corticospinal tract, which originate in the sensory cerebral cortex, end in the dorsal column nuclei and may modify their function, thus acting to filter incoming sensory messages.
The descending spinal tract of V has its cell bodies, representing all three divisions of this tract, in the trigeminal ganglion. The fibers of the tract convey pain, temperature, and crude touch sensations from the face to the first relay station, the spinal nucleus of V, or pars caudalis. The mandibular division is represented dorsally in the nucleus, and the ophthalmic division is represented ventrally. A second-order pathway arises from the cells in the spinal nucleus and then crosses and ascends to end in the thalamus.
The medial longitudinal fasciculus is an important pathway involved with control of gaze and head movements. It descends into the cervical cord. The medial longitudinal fasciculus arises in the vestibular nuclei and carries vestibular influences downward (see Fig 17–2). More rostrally in the pons, the medial longitudinal fasciculus carries projections rostrally from the vestibular nuclei to the abducens, trochlear, and oculomotor nuclei and from the lateral gaze center in the pons to the oculomotor nuclei (see Fig 8–7).
The tectospinal tract carries descending axons from the superior colliculus in the midbrain to the cervical spinal cord. It relays impulses controlling neck and trunk movements in response to visual stimuli.
Cranial Nerve Nuclei
The hypoglossal nucleus, the dorsal motor nucleus of the vagus, and the solitary tract and nucleus are found in the medulla, grouped around the central canal; in the open medulla, these nuclei lie below the fourth ventricle (Fig 7–7C). The hypoglossal nucleus, which is homologous to the anterior horn nucleus in the cord, sends its fibers ventrally between the pyramid and inferior olivary nucleus to exit as nerve XII. This nerve innervates all the tongue muscles.
The dorsal motor nucleus of X is a preganglionic parasympathetic nucleus that sends its fibers laterally into nerves IX and X. It controls parasympathetic tone in the heart, lungs, and abdominal viscera. The superior salivatory nucleus, located just rostral to the dorsal motor nucleus, gives rise to parasympathetic axons that project in nerve VII, via the submandibular and pterygopalatine ganglia, to the submandibular and sublingual glands and the lacrimal apparatus. This nucleus controls salivary secretion and lacrimation.
The ill-defined ambiguus nucleus gives rise to the branchial efferent axons in nerves IX and X. It controls swallowing and vocalization.
The solitary nucleus (still called the nucleus solitarius in some books) is an elongated sensory nucleus in the medulla that receives axons from nerves VII, IX, and X. It is located adjacent to the solitary tract, which contains the terminating axons of these nerves. The rostral part of the solitary nucleus is sometimes referred to as the gustatory nucleus. The solitary nucleus conveys information about taste and visceral sensations. Secondary fibers ascend from the solitary nucleus to the ventroposteromedial (VPM) nucleus in the thalamus, which projects, in turn, to the cortical area for taste (area 43, located near the operculum).
The four vestibular nuclei—superior, inferior (or spinal), medial, and lateral—are found under the floor of the fourth ventricle, partly in the open medulla and partly in the pons. The ventral and dorsal cochlear nuclei are relay nuclei for fibers that arise in the spiral ganglion of the cochlea. The pathways of the vestibular and cochlear nuclei are discussed in Chapters 16 and 17.
Inferior Cerebellar Peduncle
A peduncle is a stalk-like bundle of nerve fibers containing one or more axon tracts. The inferior cerebellar peduncle is formed in the open medulla from several components: the cuneocerebellar and the dorsal spinocerebellar tracts, fibers from the lateral reticular nucleus, olivocerebellar fibers from the contralateral inferior olivary nucleus, fibers from the vestibular division of nerve VIII, and fibers that arise in the vestibular nuclei. All fibers are afferent to the cerebellum.
PONS
Many pathways to and from the medulla and several spinal cord tracts are identifiable in cross sections of the pons (Figs 7–7D and E).
Basis Pontis
The base of the pons (basis pontis) contains three components: fiber bundles of the corticospinal tracts, pontine nuclei that have received input from the cerebral cortex by way of the corticopontine pathway, and pontocerebellar fibers from the pontine nuclei, which cross and project to most of the neocerebellum by way of the large middle cerebellar peduncle. Along the midline of the pons and part of the medulla lie the raphe nuclei. Serotonin-containing neurons in these nuclei project widely to the cortex and hippocampus, basal ganglia, thalamus, cerebellum, and spinal cord. These cells are important in controlling the level of arousal and modulate the sleep-wake cycle. They also modulate sensory input, particularly for pain.
Pontine Tegmentum
The tegmentum of the pons is more complex than the base. The lower pons contains the nucleus of nerve VI (abducens nucleus) and the nuclei of nerve VII (the facial, superior salivatory, and gustatory nuclei). The branchial motor component of the facial nerve loops medially around the nucleus of nerve VI. The upper half of the pons harbors the main sensory nuclei of nerve V (Figs 7–7E and 7–8). The medial lemniscus assumes a different position (lower body, medial; upper body, lateral), and the spinothalamic tract courses even more laterally as it travels through the pons.
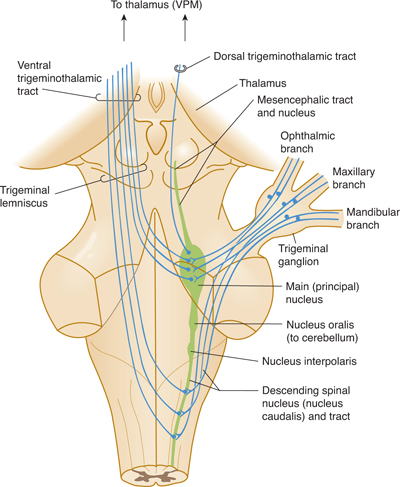
FIGURE 7–8 Schematic drawing of the trigeminal system.
Middle Cerebellar Peduncle
The middle cerebellar peduncle is the largest of the three cerebellar peduncles. It contains fibers that arise from the contralateral basis pontis and end in the cerebellar hemisphere.
Auditory Pathways
The auditory system from the cochlear nuclei in the pontomedullary junction includes fibers that ascend ipsilaterally in the lateral lemniscus (see Chapter 16). It also includes crossing fibers (the trapezoid body) that ascend in the opposite lateral lemniscus. A small superior olivary nucleus sends fibers into the cochlear division of nerve VIII as the olivocochlear bundle (see Fig 7–7D); this pathway modifies the sensory input from the organ of Corti in the cochlea.
Trigeminal System
The three divisions of the trigeminal nerve (nerve V; see Figs 7–7D and E and Fig 7–8) all project to the brain stem. Fine touch function is relayed by the main sensory nucleus; pain and temperature are relayed into the descending spinal tract of V; and proprioceptive fibers form a mesencephalic tract and nucleus in the midbrain. The second-order neurons from the main sensory nucleus cross and ascend to the thalamus. The descending spinal tract of V sends fibers to the pars caudalis (the spinal nucleus in the medulla), the pars interpolaris (a link between trigeminal afferent components and the cerebellum), and the pars oralis. The masticatory nucleus, which is medial to the main sensory nucleus, sends branchial efferent fibers into the mandibular division of nerve V to innervate most of the muscles of mastication and the tensor tympani of the middle ear.
MIDBRAIN
The midbrain forms a transition (and fiber conduit) to the cerebrum (see Figs 1–2 and 7–9). It also contains a number of important cell groups, including several cranial nerve nuclei.
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


