13 The brainstem and reticular formation
 Clinical cases for thought
Clinical cases for thoughtCase 13.2
Questions
• 13.2.1 Discuss the difference between upper and lower motor neuron lesions in general.
• 13.2.2 Discuss the different clinical signs when differentiating between an upper and a lower motor neuron lesion involving the facial nerve.
• 13.2.3 Discuss six differential diagnostic considerations for this gentleman.
Anatomy of the brainstem
The brainstem is composed of the following anatomical areas (Figs 13.1, 13.2, and 13.3):
1. Midbrain or mesencephalon is contained between the cerebrum and the pons in an area which measures approximately 2.5 cm long;
2. Pons is contained between the midbrain and the medulla;
3. Medulla oblongata extends from the base of the pons to the first pair of cervical nerves. Caudally, the medulla is continuous with the medulla spinalis or the spinal cord.
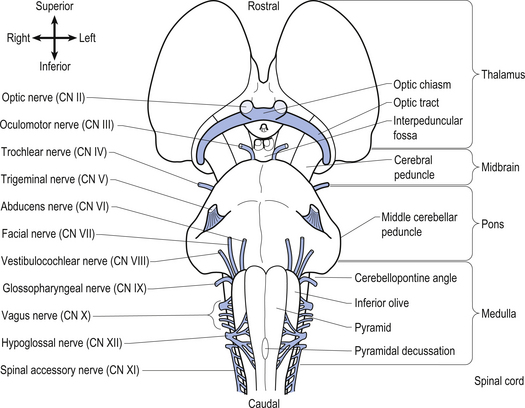

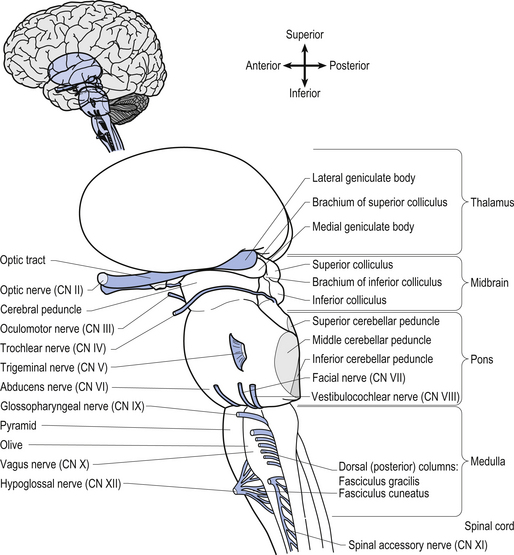
We will approach the description of the brainstem by outlining the structures observed at various levels in cross-sectional dissections.
 Quick facts 13.1
Quick facts 13.1Input from primary afferents and their collaterals results in the following
1. Monosynaptic reflex involving the alpha motor neurons
4. Inhibition of contralateral homologues
5. Excitation of antagonist of contralateral homologues
7. Excitation of granular layer of the cerebellum, which leads to increased contralateral cortical integration via excitation of contralateral thalamus.
Mesencephalon
The structures of the mesencephalon can be observed in a cross-section of midbrain at the superior colliculus and the inferior colliculus (Figs 13.4 and 13.5).
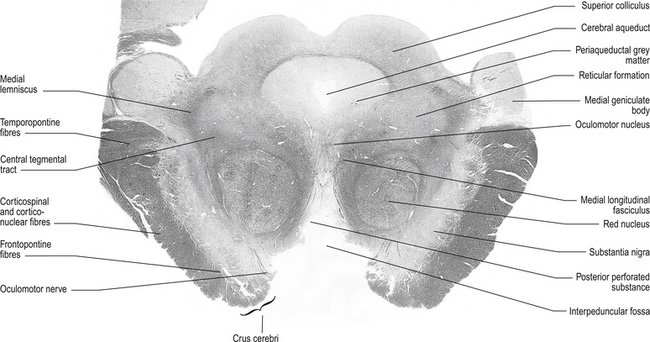
Figure 13.4 A cross-sectional view of the mesencephalon at the superior colliculus.
(from Standring 2008 Gray’s Anatomy 40th edn. Edinburgh, Elsevier, with permission)
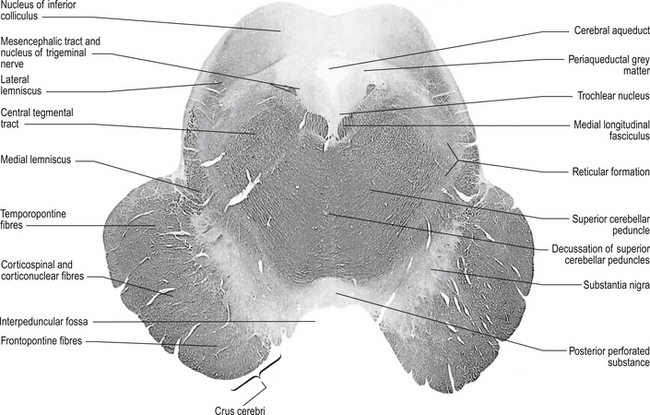
Figure 13.5 A cross-sectional view of the mesencephalon at the inferior colliculus.
(from Standring 2008 Gray’s Anatomy 40th edn. Edinburgh, Elsevier, with permission)
 Quick facts 13.2
Quick facts 13.2Tectum
The tectum comprises the ‘roof’ or dorsal portion of the midbrain and contains the corpora quadrigemina, which includes the superior and inferior colliculi and all the substances that lie dorsal to the cerebral aqueduct in the midbrain area of the neuraxis. The superior colliculi are involved with visual reflexes, and project to the lateral geniculate bodies of the thalamus. The inferior colliculi are involved with reflexes associated with sound, and project to the medial geniculate bodies of the thalamus.
Red nucleus
These bilateral structures are ovoid groups of nuclei composed of two different cell types, the magnocellular and parvocellular groups of neurons. The magnocellular neurons are large, multipolar cells located in the caudal area of the red nuclear mass. These neurons receive bilateral projections both from sensorimotor cortical areas via the corticorubral tracts and from collaterals via the corticospinal tracts. The cortical projections and their target neurons in the red nucleus are somatotopically organised. Axon projections from the magnocellular neurons form the rubrospinal tracts, which cross in the brainstem and project in a somatotopically organised fashion, mainly to the interneurons of the intermediate grey areas of the spinal cord. Some rubrospinal fibres terminate directly on ventral horn motor neurons as well. Some axons that form the rubrospinal tracts terminate on neurons in the pontomedullary reticular formation and the motor nuclei of various cranial nerves, forming the rubroreticular system and the rubrobulbar tracts, respectively (Brown 1974). Reciprocal, bilateral projections to the superior colliculi are also present and form the rubrotectal tracts (Fig. 13.6). The rubrospinal and corticospinal tracts form the lateral motor system of the spinal cord. The medial motor system is composed of the reticulospinal and vestibulospinal tracts.
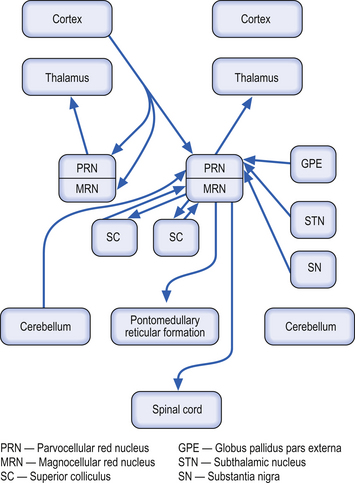
The parvocellular neurons of the red nucleus are small pyramidal- and spherical-shaped neurons, mostly located in the rostral areas of the red nuclear mass. These neurons receive projections from the dentate nucleus of the contralateral cerebellum, and from the ipsilateral globus pallidus pars externa, substantia nigra, and subthalamic nuclei. These neurons project to the ipsilateral thalamus (Fig. 13.6).
Medial longitudinal fasciculus (MLF)
This structure is a highly myelinated axon tract that descends from the interstitial nucleus of Cajal in the lateral wall of the third ventricle through the midbrain, pons, and medulla to the spinal cord where it becomes continuous with the anterior intersegmental fasciculus. The MLF acts as a major communications conduit between all of the cranial nerve nuclei and all related structures including the reticular formations of the mesencephalon, pons, and medulla. The medial vestibulospinal tract axons project with the MLF to the spinal cord (Brodal et al. 1962). Other axons from the vestibular nuclei ascend in the MLF to more rostral structures including the extraocular cranial nuclei. The MLF is the first fibre tract in the developing embryo to become myelinated and probably acts as a major pathway providing stimulus to developing neurons in the early stages of embryonic development.
Medial lemnisci
The medial lemniscus is a bundle of axons that forms a triangular structure medial to the spinothalamic tract. The bundles are formed from axons of the contralateral dorsal column nuclei, the nucleus gracilis and cuneatus, which have decussated and formed the internal arcuate fibres which become continuous with the medial lemnisci. The fibres are joined by axons of the trigeminal sensory nucleus to project to the ipsilateral thalamus. Axons from the dorsal column nuclei terminate on neurons in the ventral posterior lateral nucleus of the thalamus, whereas axons from the trigeminal sensory nucleus terminate on neurons in the ventral posterior medial nucleus of the thalamus (Guyton & Hall 1996).
Pons
The structures of the pons can be observed through a cross-section at the level of the trigeminal nerves (Fig. 13.7), just superior to the cerebral peduncles.
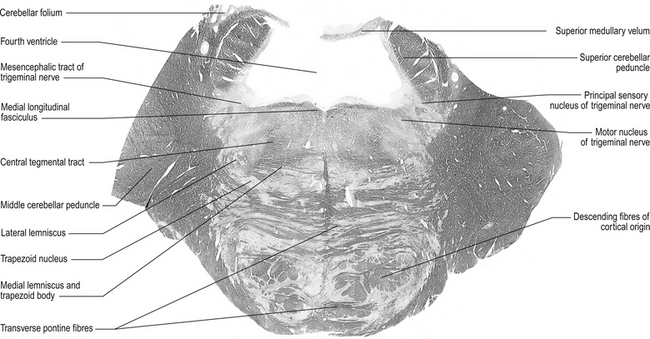
Figure 13.7 A cross-sectional view of the pons at the level of the trigeminal nerves.
(from Standring 2008 Gray’s Anatomy 40th edn. Edinburgh, Elsevier, with permission)
Superior cerebellar peduncle
 Quick facts 13.3
Quick facts 13.31. Dentatorubral and dentatothalamic fibres, both of which terminate contralaterally in the red nucleus and the thalamus, respectively;
2. Fibres of the ventral or anterior spinocerebellar tract projecting to the cerebellum from the spinal cord; and
3. Fibres of the uncinate fasciculus that contains fibres from the fastigial nucleus that will terminate in the lateral vestibular nucleus (Chusid 1982).
Lateral lemnisci
This tract carries fibres from the contralateral dorsal cochlear nucleus to the inferior colliculus.
Fourth ventricle
This cavity is bounded by the pons and medulla ventrally and by the cerebellum dorsally. It is continuous with the cerebral aqueduct above and the central canal of the medulla below and has a capacity for cerebrospinal fluid (CSF) of about 20 ml. The floor of the fourth ventricle, which is also referred to as the rhomboid fossa, is formed by the dorsal surfaces of the pons and medulla. The fourth ventricle acts as a component of the CSF system (Chusid 1982).
Medulla
Structures of the medulla can be viewed by cross-sections at:
Structures found at a cross-section at the inferior olive (fig. 13.8)
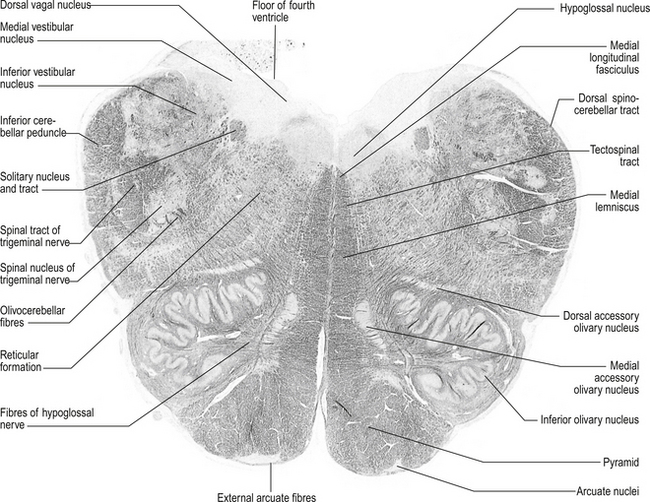
Inferior cerebellar peduncle
1. Olivocerebellar tract, which arises from neurons of the contralateral superior olivary nucleus and projects to the cerebellar hemispheres and vermis;
2. Dorsal spinocerebellar tract, which arises from neurons in the area of Clark’s column of the spinal cord to project to the interpositus nuclear region and the palaeocerebellar cortex;
3. Dorsal external arcuate fibres from the nuclei gracilis and cuneatus;
4. Ventral external arcuate fibres from the lateral reticular nuclei of the medulla; and
5. Vestibulocerebellar tract, which arises from the vestibular nuclei and projects to the flocculonodular lobe of the cerebellum.
Lateral spinothalamic tract
 Quick facts 13.4
Quick facts 13.4These tracts are located medial and anterior to the ventral spinocerebellar tracts in the ventral lateral fasciculus of the spinal cord. They are formed by axons projecting from neurons located in the contralateral dorsal horn area. These neurons project their axons via the anterior white commissure to the opposite lateral spinothalamic tract where they terminate on the ipsilateral thalamus to the tract in which they ascend.
Inferior olivary nucleus
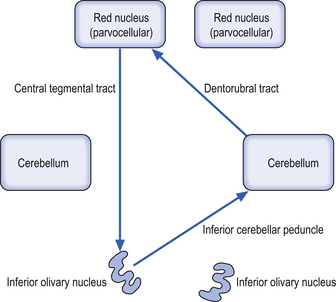
These bilateral groups of nuclei are located within the olive of the medulla. They receive projections from the cortex, from other brainstem nuclei, from the ipsilateral parvocellular red nucleus, and from the ipsilateral spinal cord. These neurons project axons, referred to as climbing fibres, via the inferior cerebellar peduncles to all areas of the cerebellar cortex. These structures are part of a group of projections and nuclei referred to as the inferior olivary nuclear complex, which forms a complicated loop from the cerebellar cortex to the dentate nucleus to the contralateral red nucleus to the inferior olivary nucleus and back to the contralateral cerebellar cortex (Fig. 13.9)
Solitary tract nucleus
These nuclei are also referred to as the nuclei of the tractus solitarius (NTS). These nuclei are located ventrolaterally to the motor nuclei of the vagus nerve and run the full length of the medulla. All cranial visceral afferent nerves project to the NTS. The rostral portion of the NTS, which is referred to as the gustatory nucleus, receives projections from the special visceral afferent nerves (CN VII, CN IX, CN X) for taste. The caudal portion of the NTS, which is referred to as the cardiorespiratory nucleus, receives projections from the general visceral afferent fibres of CN IX and CN X.
Structures found in a cross-section at the lemniscal decussation (fig. 13.10)
Fasciculus gracilis
These bilateral tracts run dorsally in the medulla medial to the cuneate fasciculi. They are formed by the axons of the dorsal root ganglion cells that detect proprioception and touch in the lower limbs and trunk, usually below the T6 level. The axons enter the spinal cord via the dorsal root and ascend ipsilaterally in the fasciculus gracilis to the gracile nucleus.
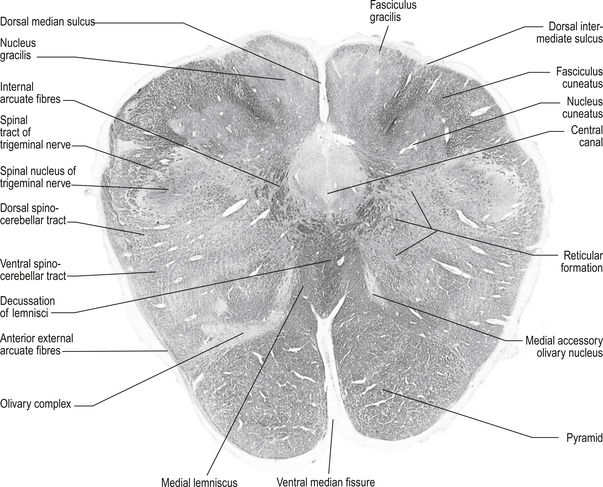
Structures found at a cross-section through pyramidal decussation (fig. 13.11)
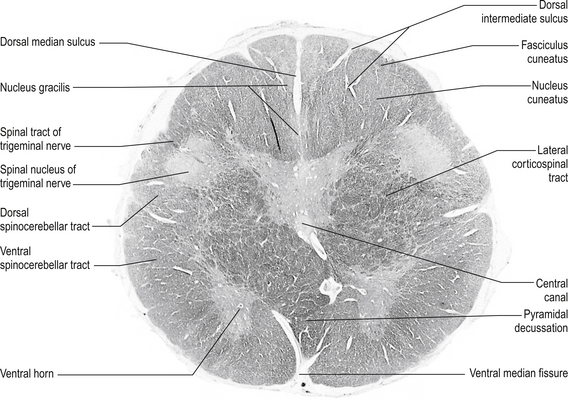
Anterior corticospinal tracts
These bilateral tracts are contained in the pyramids of the medulla until the decussation of the pyramids where they do not decussate but continue ipsilaterally in the ventral medial portion of the ventral fasciculus of the spinal cord. These tracts relay motor information to the muscles of the trunk, spine, and proximal limbs.
The reticular formation (RF)
The reticular formation receives little attention in traditional neurology textbooks. It is an area that spans all levels of the brainstem, from the thalamus to the spinal cord, and is responsible for integrating information from the brain and periphery and linking sensory, motor, and autonomic nuclei of the brainstem. The reticular formation therefore mediates complex reflexes and functions such as eye movements, posture, feeding, breathing, homeostasis, arousal, sleep, control of vasomotor tone and cardiac output, and pain. The reticular formation is composed of continuous groups of neurons interconnected via polysynaptic pathways that can be both crossed and uncrossed in nature. The RF receives projections from virtually all sensory modalities and projects to all areas of the neuraxis including direct projections to the cortex (Webster 1978).
 Quick facts 13.5
Quick facts 13.5 Quick facts 13.6
Quick facts 13.6Afferent projections to the RF include:
Efferent projections from the RF include:
• Projections to autonomic nuclei and intermediolateral (IML) column;
• Direct projections to the thalamus and cortex; and
• Direct projections to other areas of the reticular system.




