7 The spinal cord and peripheral nerves
 Clinical cases for thought
Clinical cases for thoughtCase 7.1
Questions
• 7.1.1 Describe the neural pathways involved in transmitting this girl’s pain from her leg to her brain.
• 7.1.2 Suggest a list of at least five differential diagnostic possibilities for this girl’s presentation.
• 7.1.3 Describe the presentation of CRPS and discuss the similarities and differences with this case.
Anatomy of the spinal cord
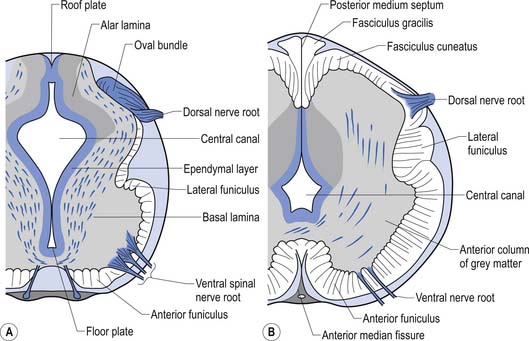
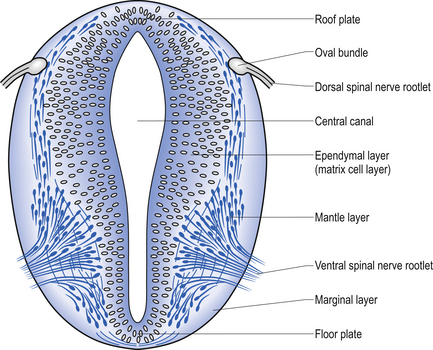
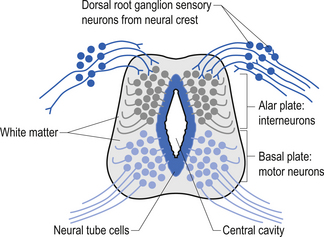
The spinal cord develops as a contiguous structure with the rest of the neuraxis, arising from the ventricular layer of ependymal cells and maintains the basic dorsal and ventral segregation of sensory and motor function as the brainstem (see Chapter 2). The result of this is that most afferent (sensory) information arrives in the dorsal aspects of the cord and the efferent (motor) information exits from the ventral aspects of the cord (see Fig. 2.9A). As the spinal cord matures during embryonic development the dorsal/ventral segregation becomes more defined and by about 3 months post-conception two discrete cellular areas can be determined: the alar lamina, which is located dorsally and contains the neurons that will receive the afferent (sensory) information, and the basal lamina, which is located ventrally and contains the neurons that will supply the efferent (motor) outflow from the spinal cord (Fig. 7.1).
 Quick facts 7.1
Quick facts 7.1The spinal cord has the following functions
1. Is the final common pathway for the somatomotor system.
2. Conveys somatosensory information from the body to higher centres.
3. Contains preganglionic ANS neurons under segmental/suprasegmental control.
4. Mediates spinal and segmental reflexes.
5. Contains central pattern generators for rhythmic movement gait and posture maintenance.
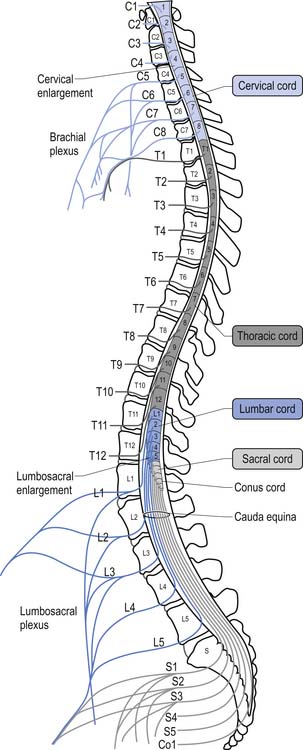
For the first 3 months of embryonic development the spinal cord and the vertebral column develop at the same pace and are roughly equal in length. During the rest of embryonic development the vertebra column grows in size faster than the spinal cord, resulting in the spinal cord terminating about two-thirds of the way down the vertebral column (Chusid 1982). The length of the spinal cord, which is usually between 42 and 45 cm, can show significant variation between individuals with the end result affecting the level of termination of the spinal cord (Barson 1970). The variation in the termination of the spinal cord can range from the lower third of the twelfth vertebra to the disc between the second and third lumbar vertebra (Jit & Charnalia 1959). The spinal cord terminates by converging into a cylindrical funnel-shaped structure referred to as the conus medullaris, from the distal end of which extends a thin filament, the filum terminale, to its attachment on the first coccygeal segment. The spinal nerve roots radiating from the spinal cord and the dorsal root ganglion neuron’s central projections form a structure referred to as the cauda equina as they traverse the distance, through the spinal canal, between the spinal cord termination point and their exit vertebral foramina in the spinal column (Fig. 7.2).
The volume of the spinal cord is dependent on the number of neurons and axons that it contains at any one point. Because of the increase in afferent input and efferent output that occurs at the level of the cervical and lumbar cord levels, due to the innervation of the arms and legs, the spinal cord expands in circumference, resulting in the cervical and lumbar enlargements.
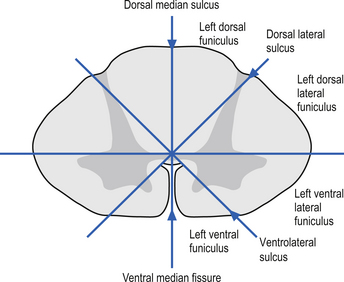
On cross-sectional views of the spinal cord, the dorsal or posterior median sulcus, which is continuous with a projection of connective tissue that penetrates the posterior aspect of the cord, the dorsal median septum, symmetrically divides the dorsal cord into two halves. Ventrally, the ventral median fissure performs a similar function so that a line connecting it and the dorsal median sulcus effectively divides the spinal cord into left and right symmetrical halves. It is convenient to divide the white matter of the spinal cord into regions referred to as funiculi, so that each half of the spinal cord contains a dorsal or posterior funiculus, a posterior lateral and anterior lateral funiculus, and a ventral or anterior funiculus (Fig. 7.3). In the mature spinal cord the embryonic alar and basal plates, with a few exceptions, maintain their distribution of sensory and motor segregation. These areas can be outlined quite accurately by the funicular divisions just described (Fig. 7.4).
The grey matter of the spinal cord is composed of a high proportion of neurons, neuroglia, and blood vessels
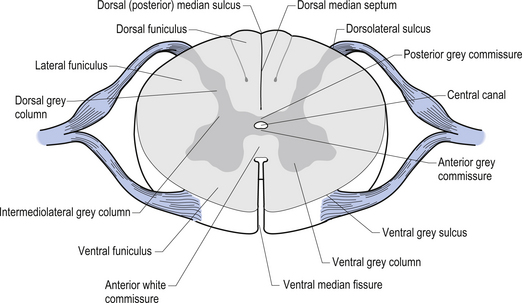
Centrally the butterfly-shaped grey matter of the cord is divided in the midline by the central canal. The grey matter passing dorsally to the central canal is referred to as the posterior grey commissure and the grey matter passing ventrally to the central canal is referred to as the anterior grey commissure. Arising from the area of the ventral lateral sulci are the ventral roots of the spinal cord, which just as they exit the vertebral foramina combine with the afferent axons of the dorsal root ganglion neurons as they enter the vertebral foramina to form the root of the spinal nerve. Entering the spinal cord at the dorsal lateral sulcus are the sensory dorsal roots, completing their journey to the cord from the dorsal root ganglion cells (Fig. 7.5). The areas of grey matter that give rise to or receive the afferent and efferent input resemble the shape of a horn and are thus termed the anterior and posterior horns. The anterior horn does not extend through the anterior funiculus and reach the surface of the cord. The posterior horn projects much more deeply into the dorsal funiculus and except for a small band of translucent neurons, the substantia gelatinosa, it would extend to the posterior surface of the cord. A small angular projection from the intermediate areas of the cord forms the lateral horn of grey matter that only occurs between the levels of the first thoracic to the second lumbar segment. This lateral outgrowth of grey matter houses many of the presynaptic neurons of the sympathetic nervous system.
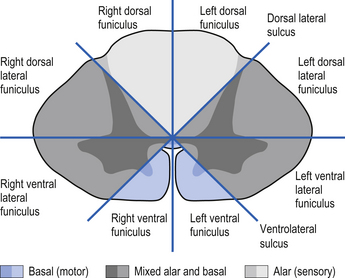
Figure 7.5 The various structures and nerve fibre pathways in a cross-sectional view of the spinal cord.
The neurons of the grey matter form a complex intermingled array involving multiple synaptic connections with many of the axons crossing the midline via the anterior and posterior commissures. Some of the neurons are intrasegmental and their axons and dendrites remain within the same segment of the spinal cord as the neuron soma. Other neurons are intersegmental and their axons and dendrites spread over many segments both rostrally and caudally. In many parts of the neuraxis groups of neurons, usually with a related functional activity, cluster together into nuclei or, when large enough, ganglia. Several nuclei have been identified in the grey matter of the spinal cord. The most predominant neurons in the ventral grey areas are the large multipolar neurons whose axons emerge from the spinal cord to form the anterior horn, and contribute to the spinal nerves, to ultimately innervate the skeletal muscles of the body. These neurons are also referred to as alpha-efferents or alpha motor neurons. Also present in large numbers in the anterior horn are slightly smaller neurons whose axons supply the intrafusal fibres of the muscle spindle called gamma-efferents or gamma motor neurons (Fig. 7.6).
The neuron groupings in the posterior horns involve four main nuclei, two of which extend through the length of the cord and two that are present only at selective levels of the cord. The substantia gelatinosa of Rolando extends throughout the cord and composes the extreme tip of the dorsal horn. These neurons are involved with signal processing of afferent information from the dorsal root ganglion neurons and are thought to play an essential role in the initial processing of pain due to extensive connections with incoming axons destined to form the spinothalamic tracts (Fig. 7.6).
 Quick facts 7.2
Quick facts 7.2Spinal cord development
• Medulla spinalis extends from the upper border of atlas to the conus medullaris opposite the L1–L2 disc.
• Filum terminale extends to the tip of the coccyx.
• Cord shows cervical and lumbar enlargements.
• In early embryonic development the cord is as long as the vertebral canal but as development proceeds it lags behind the vertebral column.
 Quick facts 7.3
Quick facts 7.3Internal structure of the spinal cord
• Spinal cord consists of a core of neuropil (grey matter) surrounded by an outer axon fibre layer, the white matter.
• The white matter decreases in proportion as the spinal cord lengthens except at the cervical and lumbar enlargements.
• Grey matter is composed of neuron cell bodies, dendrites, and efferent and afferent axons of the neurons.
A second nuclear group that extends throughout the spinal cord is located ventral to the substantia gelatinosa and is referred to as the dorsal funicular group or the nucleus proprius (Fig. 7.6). Lying ventral to the nucleus proprius in the basal region of the dorsal horn and extending from the eighth cervical region of the cord to the fourth lumbar region of the cord is the nucleus dorsalis or Clark’s nucleus (Fig. 7.6). Finally, a small group of nuclei known as the visceral grey area, or nucleus centrobasalis, is present only in the lower cervical and lumbosacral segments of the cord.
Rexed’s laminae can be used to classify functional aspects of grey matter
In the 1950s and early 1960s an architectural scheme was developed to classify the structure of the spinal cord, based on the cytological features of the neurons in different regions of the grey substance. It consists of nine laminae (I–IX) that extend throughout the cord, roughly paralleling the dorsal and ventral columns of the grey substance, and a tenth region (lamina X) that surrounds the central canal (Rexed 1964) (Fig. 7.8).
 Quick facts 7.4
Quick facts 7.4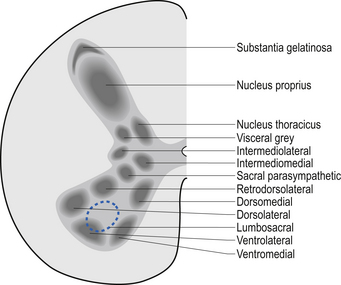
A brief description of the functional characteristics of these laminae follows.
Laminae I–IV
These areas are considered the main receiving junctions for primary afferent information. This region is characterised by complex multisynaptic networks of both intra- and intersegmental neurons. Many of the pathways cross the midline of the cord and ascend or descend in contralateral tracts (Fig. 7.9).
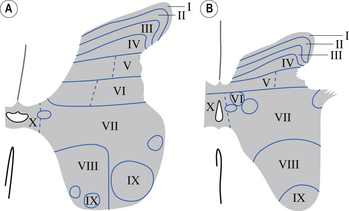
Laminae V and VI
 Quick facts 7.5
Quick facts 7.5Various rexed laminae can be referred to by more than one name
| Laminae | Names |
|---|---|
| I | Lamina marginalis |
| Layer of Waldeyer | |
| II and III | Substantia gelatinosa |
| III and IV | Nucleus proprius |
| V | Deiter’s nucleus |
| VII | Clark’s column |
| Thoracic nucleus | |
| X | Substantia gelatinosa centralis |
These areas receive proprioceptive primary afferent information as well as descending collateral input from axons of the corticospinal and reticulospinal tracts. This area is probably very involved in multimodal integration and regulation of movement (Fig. 7.9).
Lamina VII
The medial aspect of this area contains numerous complex networks of connections between propriospinal neurons. This area is probably involved in the integration of the complex propriospinal reflex network of the spinal cord concerned with both movement and autonomic function (Fig. 7.9).
Lamina VIII
This area receives collateral projections from adjacent laminae, medial longitudinal fasciculus, and vestibulospinal and reticulospinal tracts and profuse projections from the contralateral lamina VIII region. Output of this area influences both ipsilateral and contralateral neuron pools through both direct projections to the alpha motor neurons and projections to the gamma motor neuron pools (Fig. 7.9).
The white matter of the spinal cord is composed of axon fibre tracts
Axon fibre tracts of the dorsal funiculus
The dorsal columns are composed of the medially located fasciculus gracilis and the more laterally located fasciculus cuneatus (Figs 7.10, 7.11, 7.12). These pathways transport information from receptors in the periphery about fine and discriminative touch, conscious proprioception, pressure, two-point discrimination, and vibration sense. The primary afferent axons enter the spinal cord grey matter through the dorsal horn and synapse on the neurons in laminae V and VI. The secondary afferents ascend in the ipsilateral dorsal columns. Information from the lower limb and trunk is carried in the gracile funiculus and information from the upper limb and hand by the cuneate funiculus and synapse ipsilaterally in the gracile and cuneate nuclei of the caudal medulla.
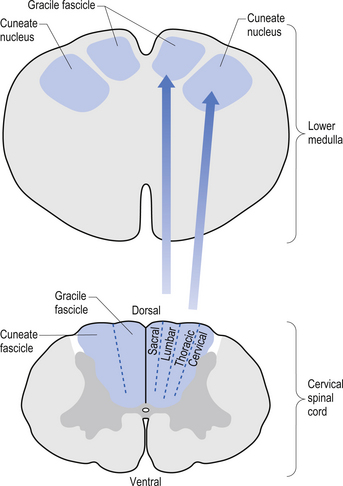
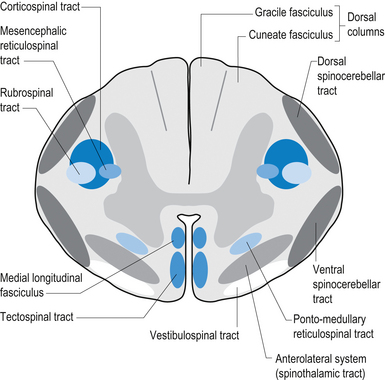
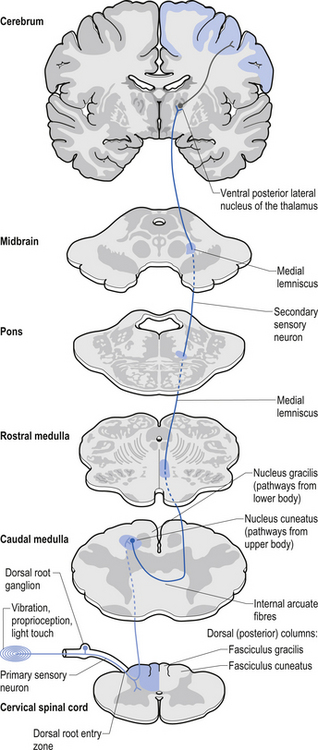
Figure 7.12 The pathway of the dorsal column/medial lemniscal system through the various levels of the neuraxis.
Axon fibre tracts of the lateral funiculus
The anterolateral system contains the fibre tracts of the spinothalamic tract and some of the fibres comprising the spinoreticular and spinomesencephalic tracts. These last two pathways provide the afferent limb for neuroendocrine and limbic responses to nociception (Figs 7.11 and 7.13).
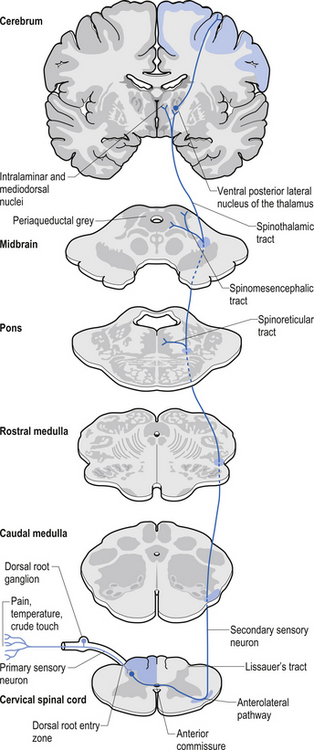
The ventral spinocerebellar pathway conveys information about the ongoing status of interneuronal pools in the spinal cord to the cerebellum. It therefore provides continuous monitoring of ascending and descending information concerning locomotion and posture. The neurons of this tract originate in laminae V–VII between L2 and S3. Their projection axons decussate to the other side so that they ascend in the contralateral anterolateral funiculus. These fibres then decussate again via the superior cerebellar peduncle to synapse on neurons in the anterior part of the ipsilateral cerebellum (see Fig. 7.11).
The primary afferent cell bodies are located in the DRG and their central processes synapse with the above-mentioned neurons near the entry level or after ascending for a short distance in the dorsal columns. Secondary afferents ascend in the ipsilateral dorsolateral funiculus, lateral to the corticospinal tracts, and enter the ipsilateral cerebellum via the inferior cerebellar peduncle. Via this pathway, the cerebellum is provided with ongoing information about joint and muscle activity in the trunk and limbs. The cuneocerebellar pathway carries the same type of information from the upper limb and cervical spine via the cuneate fasciculus of the dorsal columns (see Fig. 7.11).
Motor pathways
The anterior and lateral corticospinal tracts are important spinal tracts in the control of volitional movement. The fibre tracts are composed of axons from many different areas of the cortex as well as about 50% of their axons from unidentified areas. These tracts contain about 50% of their axon projections from the large pyramidal neurons of the primary motor cortex and association motor cortex. The lateral corticospinal tract descends in the spinal cord anterolateral to the posterior horn of grey matter and medial to the posterior spinocerebellar tract (Figs 7.11 and 7.14). It contains a large number of motor axon projections from cortical areas 1–4 and 6 to the hands, arms, legs, and feet. Its defining role is to convey motor signals to the ventral horn cells (VHCs) at the lower aspect of the cervical and lumbosacral enlargements of the spinal cord, thus controlling distal limb movements and coordinating distal and proximal muscles to achieve specific trajectories in space. Each corticomotoneuronal cell can achieve these complex goals by synapsing on interneuronal cells that communicate with whole groups of VHCs.
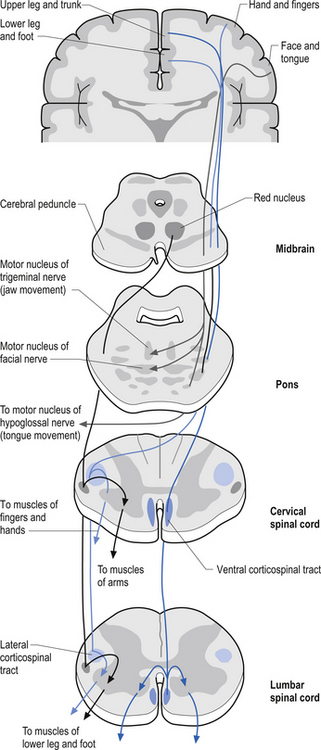
Axons from the projection neurons in the cortex descend in the internal capsule of the cerebrum through the cerebral peduncle of the mesencephalon and continue through the ventral areas of the pons until they enter the pyramids of the medulla oblongata. As the fibres descend in the medulla, about 68% of the fibres cross to form the lateral corticospinal tracts in the lateral funiculus of the contralateral side of the spinal cord; about 30% of the fibres do not cross and form the anterior corticospinal tracts in the ventral funiculus of the ipsilateral side of the spinal cord. The remaining fibres continue as uncrossed fibres of the lateral corticospinal tracts (Fulton & Sheenan 1935).
Most of the axons of both the anterior and lateral corticospinal tracts synapse on interneurons located in laminae IV to VII of the grey matter of the spinal cord (Nyberg-Hansen 1969). Most physiological evidence suggests that the majority of the corticospinal fibres of both tracts act to facilitate flexor groups of muscles and inhibit extensor groups of muscles, which is the opposite effect observed by projections of the vestibulospinal tracts. Injuries involving the corticospinal tracts affect the motor control of the peripheral muscles differently at different levels of the neuraxis. Injuries above the medulla decussation affect the contralateral peripheral muscles. Injuries below the decussation affect the peripheral muscles ipsilateral to the lesion. It must be remembered that not all of the corticospinal fibres cross the midline so that a lesion to a motor cortical area on one side of the corticospinal tract above the decussation will affect the motor control on both sides of the body to a certain extent. This contributes to the understanding of the ipsilateral pyramidal paresis observed when a decreased cortical function (hemisphericity) occurs.
 Quick facts 7.6
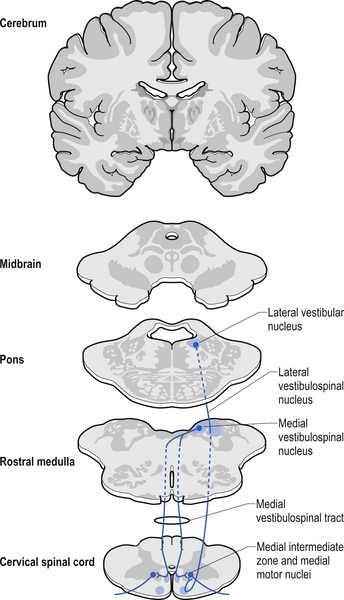
Quick facts 7.6Vestibulospinal tract
The vestibulospinal tracts, lateral and medial, descend in the ventral funiculus, and mediate reflexes (vestibulospinal) that enable an individual to maintain balance and posture despite the effect of gravity and changes in the centre of mass due to movement of the trunk, head, and limbs. The lateral segments, the lateral vestibulospinal tract, descend from the lateral vestibular nucleus uncrossed and exert modulatory effects on the ipsilateral anterior column neurons in the grey matter through the length of the cord. The medial segments of this pathway, the medial vestibulospinal tracts, arise from the medial and inferior vestibular nuclei and descend first in the medial longitudinal fasciculus before entering the vestibulospinal tracts of the cord. This pathway is both crossed and uncrossed and only projects to the cervical and thoracic levels of the cord and as such is probably only involved with upper limb and neck movements (Figs 7.11 and 7.15).
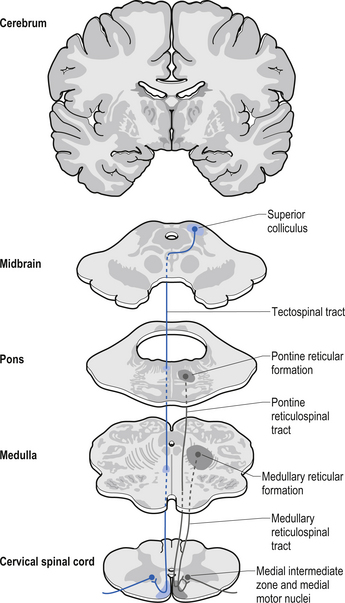
Tectospinal tract
The fibres of this tract originate from the deep layers of the contralateral superior colliculus in the dorsal midbrain (mesencephalon). The tectospinal tract crosses in the dorsal tegmental decussation of the midbrain, which is ventral to the oculomotor nucleus and the medial longitudinal fasciculus (MLF). It maintains a close relationship with the MLF until it reaches the level of the internal arcuate fibres at the decussation of the medial lemniscus. At this point it passes laterally so that it comes to lie in the ventral lateral white matter of the spinal cord. It then descends in the contralateral medial ventral funiculus of the spinal cord, synapsing on interneurons in laminae VI to VIII that communicate with the alpha and gamma motor neurons of the cervical spine (Szentagothai 1948) (Fig. 7.16).
Rubrospinal tract
The rubrospinal pathway has been rumoured to be vestigial in humans because of the evolutionary advancement of the corticospinal pathways; however, its presence in primates probably indicates it will eventually be found in humans as well and an open mind needs to prevail. It originates mainly from the large (magnocellular) neurons of the red nucleus in the rostral mesencephalon, decussates slightly more caudally, and descends just ventrally to the corticospinal fibres in the dorsolateral funiculus of the spinal cord contralaterally (Fig. 7.17).
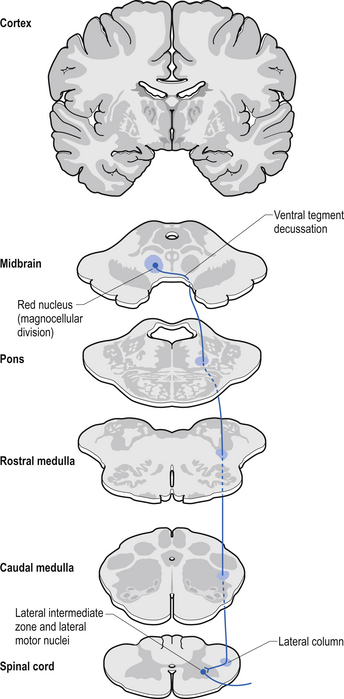
The rubrospinal tract acts much like the corticospinal tracts in that it affects enhancement flexor tone and inhibition of extensor tone, especially in the proximal limb muscles.
 Quick facts 7.7
Quick facts 7.7The history of motor function of the spinal nerve
The anterior spinal nerve roots contain only motor fibres and posterior roots only sensory fibres.
• Charles Bell’s work of 1811 contains the first reference to experimental work on the motor functions of the ventral spinal nerve without, however, establishing the sensory functions of the dorsal roots. In 1822 François Magendie definitively discovered that the anterior root is motor and that the dorsal root is sensory.
• Magendie announced that ‘section of the dorsal root abolishes sensation, section of ventral roots abolishes motor activity, and section of both roots abolishes both sensation and motor activity’.
• This discovery has been called ‘the most momentous single discovery in physiology after Harvey’. In the same volume of Journal de physiologie expérimentale et de pathologie, Magendie gave experimental proof of the Bell–Magendie Law.
• Magendie proved Bell’s Law by severing the anterior and posterior roots of spinal nerves in a litter of puppies. Stimulation of the posterior roots caused pain. Magendie sums it up: ‘Charles Bell had had, before me, but unknown to me, the idea of separately cutting the spinal roots; he likewise discovered that the anterior influences muscular contractility more than the posterior does. This is a question of priority in which I have, from the beginning, honored him. Now, as for having established that these roots have distinct properties, distinct functions, that the anterior ones control movement, and the posterior ones sensation, this discovery belongs to me’ (F. Magendie (1847) Comptes rendus hebdomadaires des séances de l’Académie des sciences, 24: 3).
Reticulospinal tract
The reticulospinal pathways can be divided into the medial or pontoreticular and lateral or medulloreticular spinal tracts. The pontoreticular neuron projections comprise the medial reticulospinal pathways and are predominately ipsilateral. They project to interneurons of laminae VII and VIII where they act to excite VHCs on the same side of origin. Some fibres do cross one or two spinal segments above their target destinations but the main modulating effects remain ipsilateral to the neurons of origin. The lateral reticulospinal pathways arise from the neurons in the medullary areas of the reticular formation and in particular from the nucleus reticularis gigantocellularis region. The projections have been found in a variety of fibre tracts in the white matter of the cord, but for the most part travel medial to the corticospinal tracts with a small tract occasionally travelling lateral to the corticospinal tracts in the lateral funiculus. In contrast with the medial reticulospinal tracts, the projections of the lateral tract are largely crossed with some ipsilateral representation (see Fig. 7.16). Projections from each half of the medullary reticular formation exert an inhibitory effect on spinal cord neurons bilaterally, probably through the activities of inhibitory interneurons (Renshaw cells) of lamina VII of the spinal cord. These projections also modulate the effects of afferent impulses arriving in these areas of the cord (Nyberg-Hansen 1965). The loss of inhibitory projections to the spinal cord from the cortex has been thought to play an important role in spasticity observed in lesions of the cord, brainstem, or cortex. However, extrapyramidal, reticulospinal inhibitory dysfunction is also thought to be an important contributing factor. The differential activation of VHC groups by reticulospinal projections (e.g. locomotor and inhibitory systems) in combination with the effects of lesions of the corticospinal projections as previously discussed leads to a characteristic weakness or ‘soft’ weakness pattern in the limbs in response to spinal cord lesions, brain damage, or hemisphericity.
Interstitiospinal tract
The fibres of this tract arise from the interstitial nucleus and descend in the medial longitudinal fasciculus. They extend into the spinal cord from the MLF into the ipsilateral fasciculus proprius, which terminates on a network of interneurons located in the dorsal horn. These interneurons are thought to participate in intersegmental coordination of various muscles.
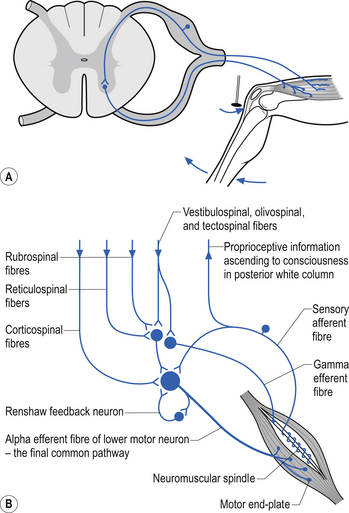
Spinal cord reflexes
Local spinal cord reflex circuits are also important in volitional movement in that descending motor pathways converge on interneurons to allow complex movement patterns to occur – i.e. corticomotoneuronal cells of the brain alter the trajectory of a limb in space by activating these reflex circuits involving agonist, antagonist, synergist, and neighbouring joint muscle groups. Feedback and feedforward mechanisms are employed by the cerebellum to assist plastic changes in the brain and spinal cord. Some stereotypical reflexes mediated by the spinal cord are state- or phase-dependent. For example, activation of Golgi tendon organs (GTOs) in the soleus and gastrocnemius muscles will trigger a different set of interneurons in the spinal cord, depending on whether the individual is in a state of locomotion or is non-ambulatory, and whether the individual is in the swing or stance phase of gait (Fig. 7.18).
Laminar organisation in the spinal cord is not complete or as accurate as previously thought
Laminar organisation in the spinal cord is not as complete or as accurate as previously thought. The traditional understanding of the laminar distribution of pathways in the white matter of the spinal cord was that the projections to and from the most distal areas of the body were more lateral in the spinal cord except in the dorsal columns where the reverse occurs. Some variability of these laminar patterns has been demonstrated; however, the general pattern in dorsal column, spinothalamic, and corticospinal tracts is important to understand from a clinical perspective.
 Quick facts 7.8
Quick facts 7.8



