In the previous chapter we saw how information is conveyed electrically from one part of a neuron to another: by passive conduction when the distances are short enough to permit it, and otherwise by means of action potentials. However, you need to keep reminding yourself that the purpose of a neuron is not to generate action potentials – or any other kind of potential – but to release transmitter in response to stimuli. We begin by looking at how neurons respond to stimuli, from the outside world or from other neurons, and then at how ultimately they secrete their neurotransmitter.
Common features of all neurons
The schematic neuron
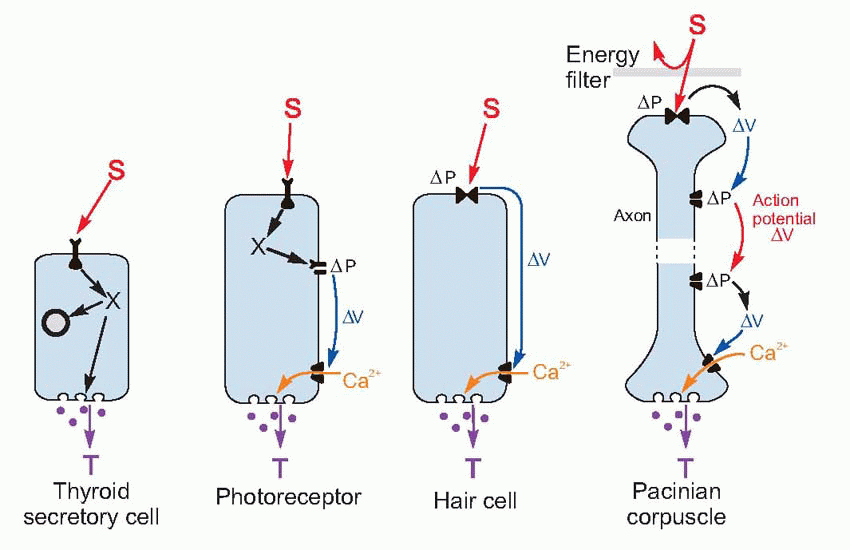
Despite the huge variety of shapes and sizes and functions of neurons, from the tiny sensory hair cells in the ear to the huge neurons that carry commands from the cerebral cortex to the bottom of the spinal cord, there is a basic ground plan for intercellular communication that applies to all of them, and indeed to endocrine cells as well.
All neurons have an output region (the terminals that release the neuron’s transmitter, T) and an input region (the dendrites, or the receptive region in the case of sensory receptors). The mechanism at the terminal end is as far as we know absolutely identical in all neurons and receptors: depolarization opens voltage-sensitive calcium channels, and the resultant rise in intracellular calcium causes exocytosis of vesicles about 50 nm across, containing the neurotransmitter that is to act on the next cell along.
In the input region, a stimulus S acts on membrane channels that open (or sometimes close). As we have seen, this action may be a direct one, or may be indirect, mediated by intracellular mechanism. Either way, these channels create changes in ionic permeability that in turn give rise to currents and potentials ΔV that in short neurons may spread passively to the terminal, or in longer ones may induce repetitive action potentials as described in Chapter 2 (p. 39). Often – for example in the hair cell receptors of the ear – the receptor and the nerve axon that joins it to the central nervous system are quite separate, the receptor cell acting on the axon by means of a chemical transmitter whose release depends on the amount of depolarization of the ending. The only fundamental difference between sensory receptors and central neurons is the nature of the stimulus that acts on the channels in the first place.
Once this basic neuronal ground plan is understood, differences between particular neurons and receptors become a matter of filling in the blanks in a rather simple table (Table 3.1). We need to know only: the type of stimulus, the type of receptor channel, what permeability change occurs, whether action potentials are used and what transmitter is finally released. What could be simpler?
The initiation of activity
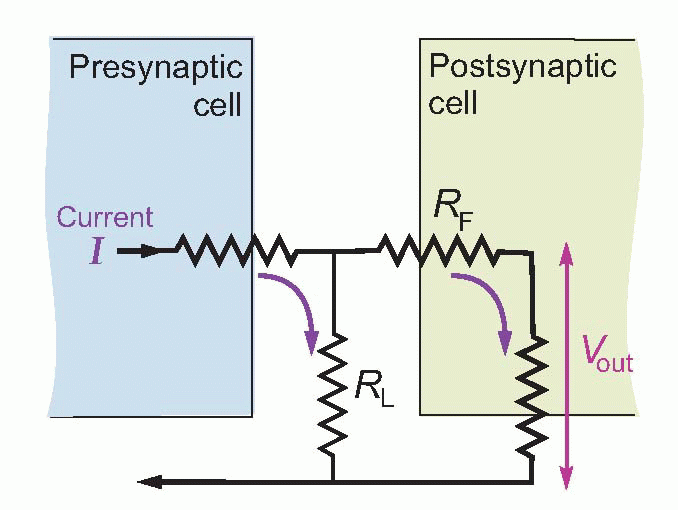
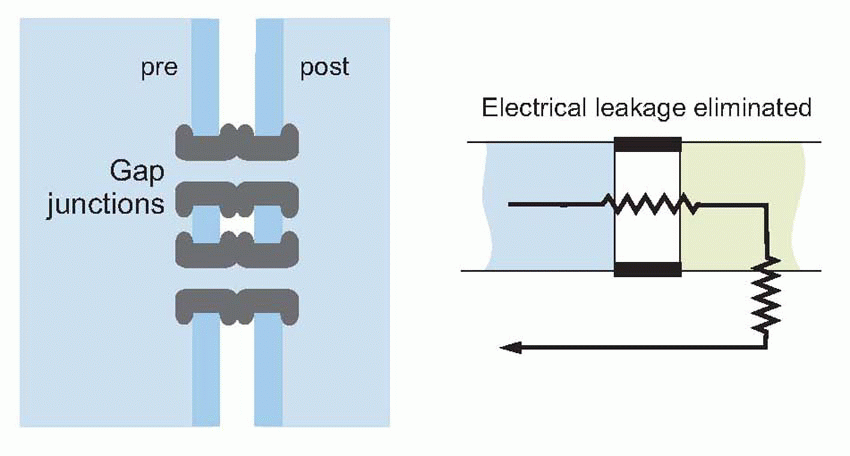
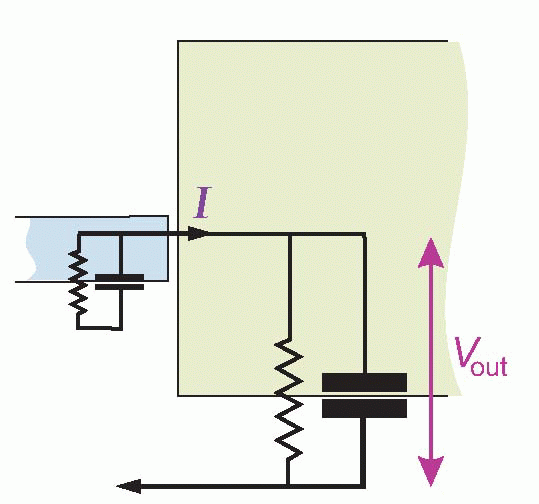
The simplest of all kinds of intercellular communication is when currents pass directly from one cell to another. But rather stringent structural conditions have to be met before this mode of synaptic transmission will work. Considering an idealized electrical synapse and its equivalent circuit, it is clear that the current I generated by the presynaptic bouton has two alternative routes: it can either cross the gap and enter the postsynaptic cell, or it can simply leak out sideways through the synaptic cleft. The greater the fraction of current that takes the former route, the greater will be the degree of electrical coupling between the two neurons, since by entering the postsynaptic cell the current will cause potential changes that may, if large enough, trigger a new action potential. More formally, if I is to create a sufficient depolarization Vout of the postsynaptic cell, this sideways leakage through RL must be small relative to the forward resistance RF. One way to minimize the sideways leakage is to use gap junctions, which enormously increase RL as well as reducing RF. A gap junction is a sealed bridge along which electrolytes can move from one cell to another without escaping to the extracellular fluid.
Table 3.1 Theme and variations: some types of neuron
Nicotinic
M2 muscarinic
Photo-receptor
Olfactory receptor
Hair cell
Pacinian corpuscle
Stimulus
ACh
ACh
Light
Chemical
Mechanical
Mechanical
Indirect?
No
Yes
Yes: cGMP
yes: cAMP, and Ca2+
No
No
ΔPNa?
+
–
+
+
+
ΔPK?
+
+
+
+
+
Action potentials?
Yes
(No)
No
Yes
No
Yes
Result
Mechanical
Mechanical
Glutamate
Glutamate?
Glutamate?
Glutamate?
A familiar example is conduction between the muscle fibres of the heart, where the currents pass through the gap junctions that form part of the intercalated discs. Electrical transmission between neurons appears to involve either casual sets of gap junctions as are found between photoreceptors in the retina, or more organized regions of contact called electrical synapses, which include gap junctions. But even if no leakage at all occurs, and the transmembrane resistance at the junction is reduced to zero, there is still no guarantee that an action potential will be able to pass successfully from one cell to another. The local currents that flow during the passage of an action potential along an axon are strongly dependent on the size of the axon. The larger it is, the greater the number of sodium channels per unit length and so the larger the active currents that can be generated. But equally, larger axons have greater capacitance and smaller transverse resistance per unit length, so the currents have to be that much larger to achieve a particular threshold level of depolarization. In other words, the currents automatically keep up with the increased requirements as the fibre’s diameter is increased.
But if we imagine a small axon whose diameter suddenly gets bigger at a particular point, or – as comes to the same thing – a small axon with a low-resistance electrical synapse joining it to a cell body of larger size, this is obviously no longer the case. Just as a burning thread may not generate enough heat to ignite a rope to which it is attached, so the current generated by the small axon may well be too weak to trigger an action potential in the larger cell. One can calculate, for example, that electrical transmission across the ordinary neuromuscular junction (NMJ) is in principle impossible even if the junction was a low-resistance one (which it is not): the impedance ratio on the two sides is much too large for the axonal currents to make any significant impression on the potential of the muscle cell. It is clear, therefore, that in such cases an extra source of amplification in addition to that provided by the action potential mechanism is needed, equivalent to soaking the knot between thread and rope in petrol.
Ligand-gated channels
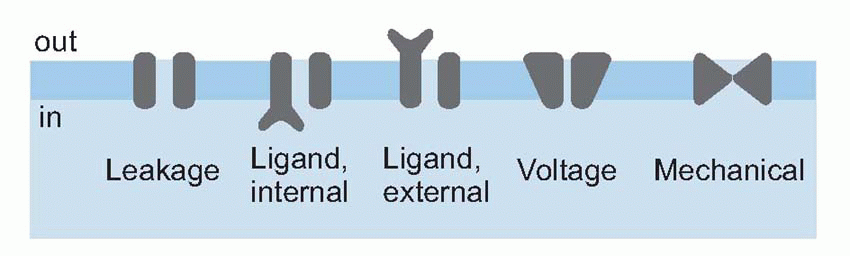
This amplification – the petrol – is provided by membrane channels sensitive not to voltage but to chemical transmitters. In general, membrane channels may be controlled by one of three things: by mechanical stimulation, by depolarization, or (in ligand-gated channels) by the arrival of particular molecules at sites that can be either on the inside or the outside of the membrane: these fundamental classes of channel can be represented symbolically as below. There is, in fact, very great similarity between the way channels in sensory receptors respond to stimuli and those in interneurons respond to transmitters. Whether it is a sensory stimulus acting on a sensory receptor, or transmitter released by one neuron acting on the postsynaptic membrane of another, what happens in every case is the opening (sometimes the closing) of particular ionic channels in the cell membrane. This leads to changes in the ionic permeability (nearly always to one or more of sodium, potassium or chloride), which must inevitably cause a change in membrane potential. This change has different names at different sites: in receptors it is called the receptor potential, at the NMJ the endplate potential, and at central synapses the postsynaptic potential; but they are all the consequence of the same underlying process. However, there are two fundamentally different ways in which a stimulus can open channels, directly or indirectly. These two modes are mediated by what are officially and inelegantly called ionotropic and metabotropic mechanisms, respectively. The basic difference between the two kinds of channel is that the direct ones are looking outward, waiting for signals to arrive from the outside world, whereas the indirect ones are looking inward, for messages that are generated within the cell itself.
Direct (ionotropic) mechanisms
One example of a direct gating mechanism is found at the muscle endplate, or NMJ. This is technically a cholinergic synapse, meaning that the transmitter is acetyl choline (ACh), and the receptors belong to the subclass of cholinergic receptors called nicotinic, because they also respond to the well-known noxious substance nicotine. Recognition of the transmitter causes opening of an unselective, short-circuiting channel, permeable to sodium and potassium. The result is to cause depolarization and the generation of an action potential in the muscle; it is discussed in more detail later (Transmission at the NMJ, p. 52). In addition, all mechanoreceptors appear to work by a direct mechanism of this kind, with the mechanical stimulus acting immediately to cause opening of the channel. Examples include the hair cells in the cochlea of your inner ear that respond to sound vibration, or the touch receptors in your skin.
Indirect (metabotropic) mechanisms
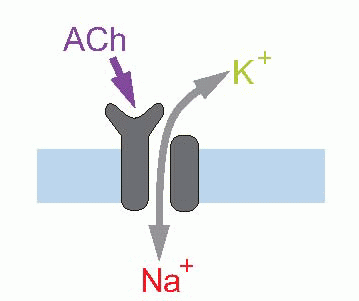
The other possibility is indirect gating; here the channel is inward-looking, responding only to chemical messages from inside the cell. The link with the outside world is provided by a second protein that straddles the membrane and responds to transmitters or stimuli in the outside world by triggering off a chemical response on the other side of the membrane, which then results in the required message being sent to the channel. This intracellular communication may involve just one intermediate, or a cascade of several of them; very often the first link in the chain is formed by a G-protein – guanosine triphosphate (GTP)-binding protein. An example of short indirect coupling is the M2-muscarinic ACh receptor, where the G-protein, activated by an ACh receptor, then acts directly on a potassium channel to cause hyperpolarization. Sometimes the G-protein activates the production of a second messenger, which may in turn either have intracellular effects (as with the β-adrenergic receptor above on the right), or again may have an effect on membrane channels. The transduction mechanism in retinal rod receptors that allows them to respond to light is a good example of a cascade of this kind: here a photolabile pigment molecule, rhodopsin, is coupled to a G-protein that activates a phosphodiesterase (PDE), that in turn results in the conversion of cyclic guanosine monophosphate (cGMP) to GMP; since cGMP opens sodium channels on the surface of the receptor cell, the effect of light is to close them, and thus to cause hyperpolarization. A similar cascade, but with cyclic adenosine monophosphate (cAMP) instead of cGMP, and further amplified by a second stage involving calcium, can be found in olfactory receptors (right, above).
These two general methods of generating permeability changes each have their advantages. Direct activation is fast and secure. Indirect activation provides for amplification (in rods, one photon can trigger the breakdown of a million or so cGMP molecules), for prolongation of effects, for control by the cell (which can intervene in the link between receptor and channel), and for intracellular effects. An example of cellular control is again in photoreceptors, where intracellular calcium modifies the sensitivity and time-course of the cGMP changes, thus effectively altering the receptor’s sensitivity to light. An obvious example of intracellular effects is the β-adrenergic response to noradrenaline, where cAMP production is again coupled via a G-protein to the receptor itself, the cAMP then having metabolic effects within the cell.
Calcium
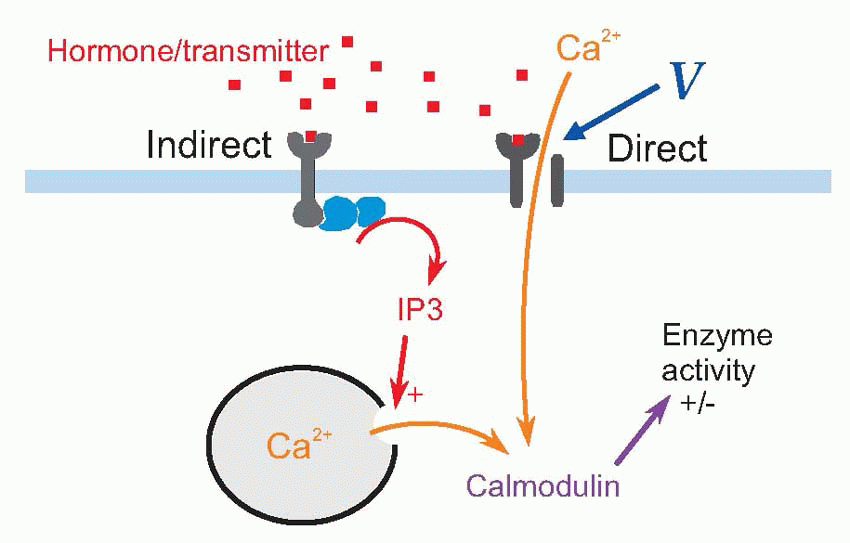
This is a good moment to say hello to calcium, an ion which is used as a form of intracellular communication by very many kinds of cells apart from neurons and muscles. The significance of calcium lies in the fact that its concentration inside cells is normally very low indeed, something of the order of 0.1 µM, because of specific calcium pumps, by its sequestration within the cell, and in some cases because of its storage in organelles that act as internal stores. Because cellular calcium concentration is effectively zero, the sudden appearance of even a tiny amount of free calcium inside is a spectacular event. Often the cell uses this as a means of telling the interior that something has happened at the membrane surface, very like ringing the cell’s doorbell. There are two ways in which this signalling can occur. Calcium can enter from outside, through channels triggered either by a transmitter or hormone or by voltage, or these signals may operate indirectly, for instance by causing the production of a second messenger such as inositol (1,3,5) triphosphate (IP3), that causes calcium to be released from the internal stores. Again and again we seeing calcium entry generating gigantic consequences (transmitter release from synaptic terminals, muscular contraction, enzyme secretion) or – as in many sensory receptors – having profound regulatory effects on the behaviour of the entire cell.
Different transmitters
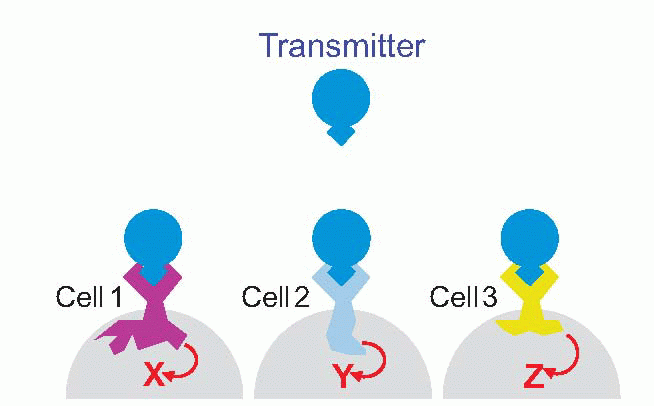
Bear in mind that the same transmitter may have quite different effects on different cells: there is no logical or necessary connection between the identity of a transmitter and what it does to the target cell – everything depends on what receptors are expressed in the target membrane. You are probably already familiar with the different effects that ACh can have in the autonomic system, mediated by nicotinic or muscarinic receptors. A particularly clear example that we come across later, in Chapter 7, is the fact that although retinal photoreceptors release only glutamate as their transmitter, the bipolar neurons on which they act may be depolarized or hyperpolarized, depending on whether they have one kind of glutamate receptor or another.
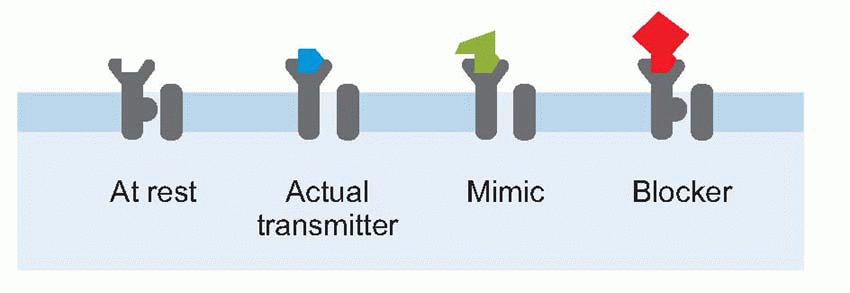
Finally, in general a receptor ‘designed’ for a particular transmitter will in general also respond to a range of other substances that may mimic the transmitter, or block it by becoming attached but refusing to budge, or in more complex ways. Many of these substances are natural poisons: many more are the artificial creations of pharmacologists, and as important in trying to elucidate how the receptor works as they are in clinical therapy.
Clinical box 3.1 Synapses as drug targets
The synapse provides one of modern medicine’s most common drug targets. Valium, for example (also known as diazepam), is one of a group of drugs called benzodiazepines used to control anxiety and seizures. The drug binds to a specific benzodiazepine binding site on the γ-aminobutyric acid (GABA)-A receptor causing the chloride channel to favour the open state. As a result, the neuron bearing the receptor is reduced in its activity. This causes a general reduction in activity in the central nervous system, which clinically manifests as relaxation or cessation of seizure activity. In overdose, predictably, patients become sedated, comatose, hypoventilate and can even die.
Indirectly, most antidepressants also act by augmenting synaptic activity. While the organic aetiology of depression is poorly understood and almost certainly multifactorial, one line of thought holds that at least part of the underlying problem is a lack of serotonin activity in the brain. This has stimulated the development of serotonin selective reuptake inhibitors (including Prozac) which inhibit the reuptake of serotonin by the presynaptic neuron: serotonin is present in the cleft for longer and activity in the postsynaptic neuron is prolonged.
Even less nuanced, Parkinson’s disease is thought to result from reduced dopaminergic transmission in the nigrostriatal part of the basal ganglia (see p. 241). A cornerstone of therapy is the oral administration of dopamine precursors to improve the symptoms of the disease.
What is astonishing is not our clever design of pharmacological agents to delicately alter neural physiology, but rather that splashing empirical, undirected drugs around the brain, little more sophisticated than throwing a can of petrol at a car, should prove to be so safe and effective!
Synaptic transmission
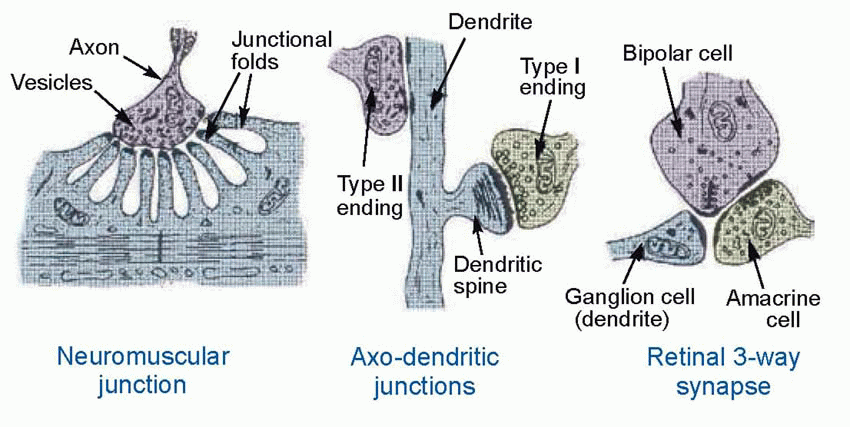
Central neurons are driven not by sensory stimuli in the outside world but by the activity of other neurons that make contact with them at specialized regions, the synapses. At a typical synapse, a branch of the afferent axon forms a swelling, the terminal bouton, the further side of which forms an enlarged area of intimate contact with the postsynaptic cell body: in the case of the neuromuscular synapse, the muscle endplate, this area is much increased by the presence of invaginating folds. In most cases there is a clear synaptic cleft between the pre- and postsynaptic membranes, typically some 20 nm wide. Transmitter is released from the presynaptic side and diffuses to the postsynaptic side, where it causes permeability changes through the various mechanisms already outlined. Anatomically, many variations on this basic pattern of axo-somatic contact can be found. Most neurons have an elaborately branched dendritic tree which is smothered in axo-dendritic synapses; dendrites may also make contact with each other in dendro-dendritic contacts; and in axo-axonic contacts one axon may act on the terminal of another and modify its transmitter release. But it is convenient to begin with the most familiar type of synapse of all, whose working is most thoroughly understood: the NMJ between motor axon and striated muscle fibre. Although the postsynaptic cell is not a neuron, the fundamental mechanism by which it is activated is closely similar to many kinds of synapses within the brain.
Transmission at the NMJ
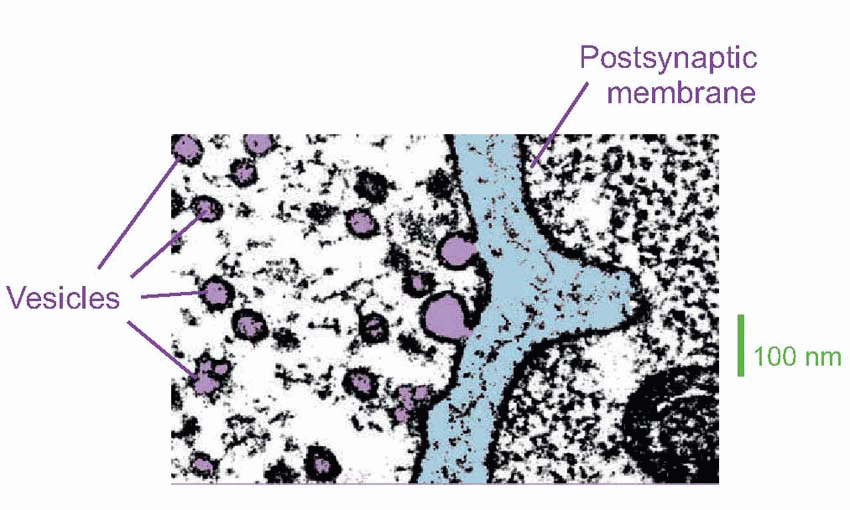
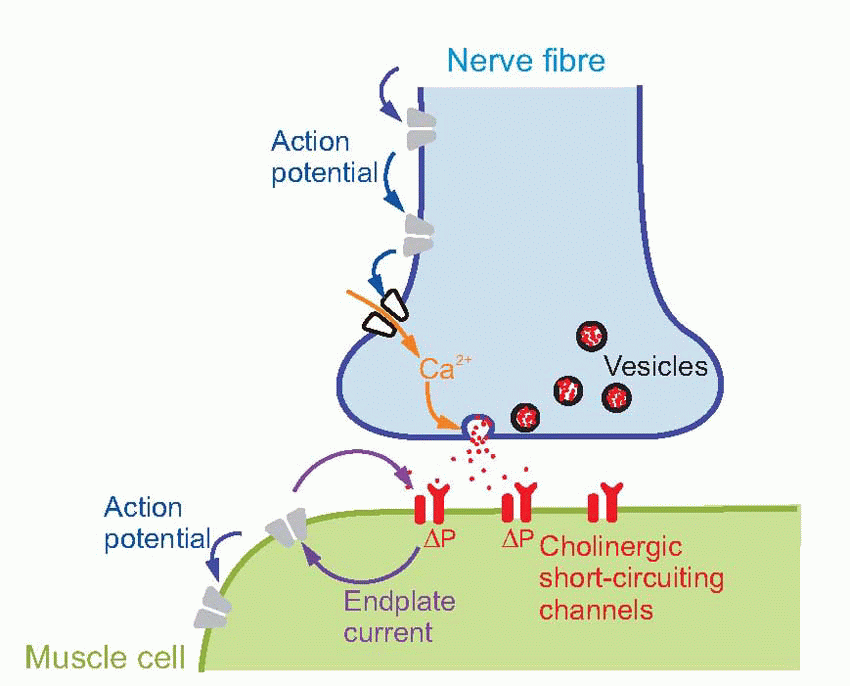
The transmitter at the NMJ is ACh. The vesicles in which it is contained are normally created by pinching off parts of the transmitter-filled Golgi apparatus in the cell body, and are then transported down microtubules in the axon to the terminal. Each is about 40-60 nm in diameter and contains some 105 molecules of ACh, and when an action potential arrives at the endplate the consequent entry of calcium triggers the release of the contents of some 200-300 of them: in the picture above of rat NMJ, you can see that a couple of vesicles have fused with the membrane to release their contents. Vesicles appear to obey a kind of all-or-nothing law in that they either empty completely into the synaptic cleft, or not at all. The transmitter thus released must diffuse across the synaptic cleft – a process that takes a millisecond at most – before it can act on the muscle cell. When it arrives there, it interacts with direct (ionotropic) channels that open to increase the permeability to sodium and potassium, thus depolarizing the membrane and initiating an action potential. How do we know all this?
The effect of ACh at the endplate
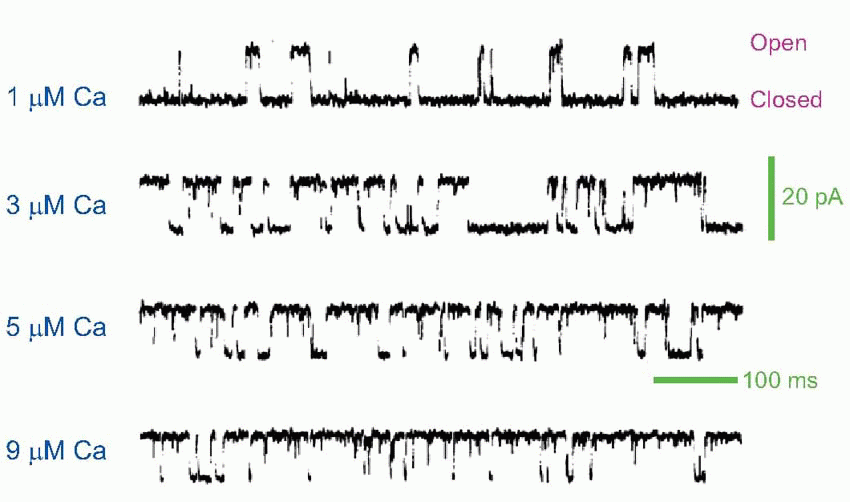
Early experimenters had to work backwards, from the changes in potential they observed to deductions about the action of ACh on the receptors. If we put a microelectrode in the muscle fibre close to the endplate, and poison the muscle with tetrodotoxin so that our observations of the primary electrical events are not obscured by any subsequent action potentials that may be generated, we find that a single action potential in the afferent nerve is associated with a characteristic electrical response in the muscle cell, called the endplate potential (EPP). Although the original action potential only lasts a millisecond or so, the EPP is relatively prolonged. Most of this prolongation is due to the capacitance of the membrane, which is first rapidly discharged and then slowly recharged to the resting potential ER. If we know the value of the appropriate time constant, we can estimate the duration of the current that flowed through the ACh-gated channels to produce the potential change. It turns out to have a time-course (blue trace below) not very different from that of the original action potential, though delayed in time by the millisecond or so of synaptic delay that is the result of diffusion across the synaptic cleft. This brief current discharges the membrane capacitance, which subsequently must recharge relatively slowly through the resting membrane resistance, giving a long tail to the resultant synaptic potential. The fact that the current hardly lasts longer than the afferent impulse is largely due to the presence at the ending of high concentrations of the enzyme cholinesterase that mops up the ACh almost as soon as it arrives. If this enzyme is blocked by an anticholinesterase such as eserine, one finds that the current flow, and hence the EPP, is enormously prolonged, leading to a depolarization block of the muscle fibre. Patch-clamping demonstrates that the channels operate in an all-or-nothing manner, being either fully open or fully closed, as can be seen in the recording from rat NMJ below – which also shows the increase in the probability of opening with increasing calcium concentration. In addition, this technique can be used to demonstrate that two ACh molecules are required to trigger the opening of one channel.
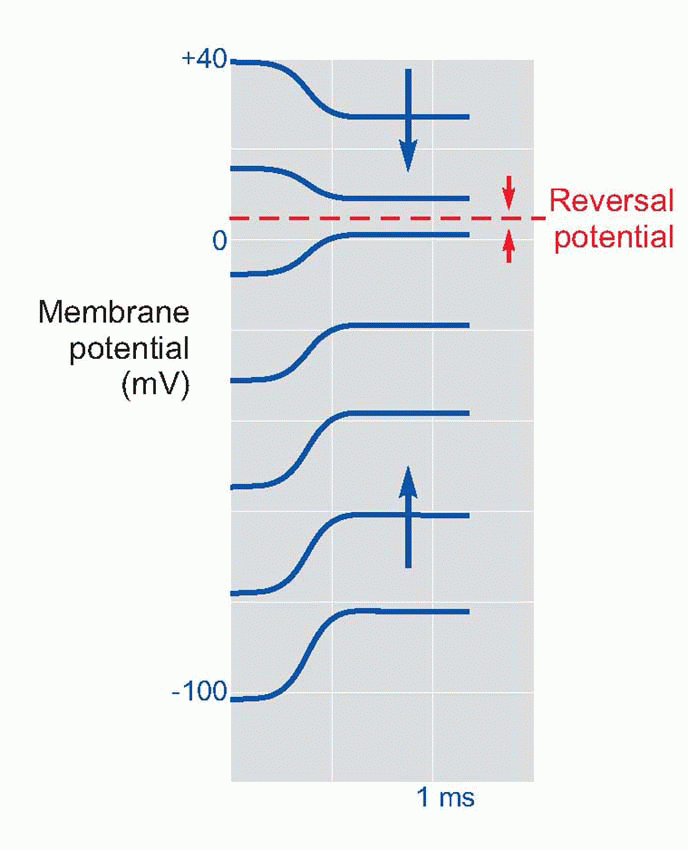
To understand what these channels do, the first thing we need to know is what ions they let through when they are open. One way to find out is to measure something called the reversal potential. This is a fundamental technique that forms a basic way of determining what permeability changes are going on in sensory receptors and in postsynaptic membranes. The principle is a simple one: since every combination of permeabilities results in some corresponding equilibrium potential Es (from the constant-field equation, p. 25), then a change in equilibrium potential implies a change in one or more of the permeabilities. The problem is that most sensory and synaptic events are short lived, so that there is no time for the membrane potential actually to settle down at its new value of Es. But the direction of its movement tells us whether the new equilibrium is above or below the resting potential; and if we have some way of setting the resting potential artificially to different levels, we can see how this influences the direction of the response. As the resting potential is made to approach Es, the response will get smaller and smaller, then reversing in sign as the resting potential passes through Es. The reversal potential, defined as the value of the resting potential at which stimulation has no effect, is simply equal to Es. Once we know Es we can make an informed guess as to what permeability change must be causing it.
In the case of the NMJ, a steady current is passed into the muscle cell in order to set the resting potential at a new artificial level, and the size of the EPP is then observed. As illustrated schematically above, the EPP gets smaller and smaller as the resting potential is reduced to near zero, and if the membrane is hyperpolarized, the EPP is reversed. The conclusion is therefore that the effect of ACh is to open channels that allow sodium as well as potassium to pass through the postsynaptic membrane, producing something like a short-circuit. Since the number of ACh molecules released by each impulse, and hence the number of channels opened, is very large, it is clear that this is the mechanism whereby the relatively small currents in the axon can trigger off the relatively enormous currents needed to initiate an action potential in the muscle cell: the source of these currents is the muscle cell itself.
The release of transmitter
The ultimate action of all neurons, whether interneurons or sensory receptors, is to release a chemical transmitter from their terminals. As far as we know, the mechanism by which this occurs is identical in every case: depolarization opens voltage-gated calcium channels, calcium enters and causes the transmitter to be released from its vesicles. The process has been most extensively studied at the NMJ because of its relatively greater size and accessibility. The details of the mechanism by which calcium causes docking of the vesicles at the synaptic membrane, and then exocytosis of their contents, are, however, still not understood.
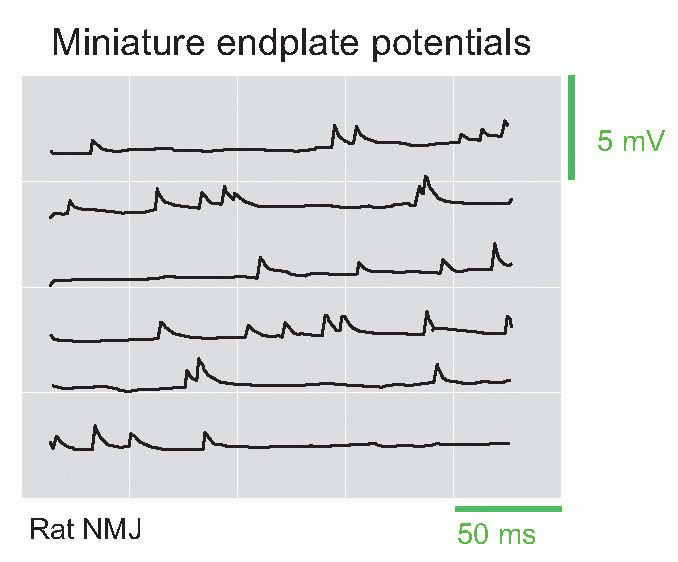
Recording from the endplate with very high sensitivity, one finds that even when the afferent fibre is not stimulated there are continual spontaneous potential changes taking the form of a random succession of miniature endplate potentials (mEPPs), having roughly the same shape as a normal evoked EPP, but about 0.2-0.3 per cent of its size. They are due to the fact that the presynaptic ending, even at rest, releases individual vesicles randomly at a very low rate. This rate of spontaneous release is strongly dependent on the resting potential across the presynaptic terminal, and if this is artificially reduced – for example by changing the external potassium concentration – the average rate of vesicle release increases sharply. By extrapolation, one can show that the size of a normal EPP is about what would be expected if the action potential simply had the effect of temporarily increasing the rate of spontaneous release of vesicles. The size of the mEPPs does not alter with different degrees of depolarization, implying that the transmitter packaged in a vesicle is either released or not released, in packets of fixed size called quanta. It is important to realize that vesicle release is a probabilistic phenomenon. At rest, the probability of a vesicle being released per unit time is very low; what happens when the terminal is depolarized and calcium enters is simply that this probability is increased. Thus the frequency of the mEPPs goes up, but their size doesn’t; mEPPs provide a convenient way of determining how certain drugs act on synaptic transmission. Some well-known poisons (for instance δ-tubocurarine, found in the arrow-poison curare, or the snake venom α-bungarotoxin) work by blocking the action of ACh on the postsynaptic membrane, whereas others (for instance botulinum toxin, found in decaying meat) operate by interfering with release from the terminal; the former affect mEPP size but not frequency, the latter frequency but not size.
Finally, the role of calcium entry into the terminal can be demonstrated by looking at the effect on quantal frequency of altering calcium concentrations outside the terminal, or of adding magnesium, which blocks its entry. The calcium inside the terminal can in addition be directly visualized by using a rather handy substance called aequorin, derived from luminescent jellyfish, that lights up in response to calcium.
The diagram above summarizes the sequence of events at the NMJ, and their relation to the three kinds of membrane channel found there: voltage-gated sodium and potassium (grey), voltage-gated calcium (white) and cholinergic ligand-gated (red). The action potential invades the terminal, channels in the presynaptic ending then open, permitting the entry of calcium, and this in turn stimulates the emptying of the vesicles into the synaptic cleft. The ACh that they contain diffuses across, reaching nicotinic receptors in the postsynaptic membrane, which respond by increasing the permeability to sodium and potassium. This generates a short-circuiting current tending to pull the membrane potential towards zero, and which in turn depolarizes the surrounding membrane sufficiently to initiate an action potential.
Clinical box 3.2 Neuromuscular blockade facilitates certain types of surgery
Even when a patient is deeply unconscious and not experiencing any pain, the muscles can be active. For example, during abdominal surgery contraction of the abdominal muscles in an effort to breathe makes suturing the abdominal wall together difficult; often the surgeon asks the anaesthetist to ‘paralyse’ the patient and let the ventilator take over the work of breathing while the surgeon completes the suturing. Similarly, during orthopaedic surgery, the trauma and irritation of cutting open a joint, whilst not ‘felt’ by the anaesthetized patient, does result in reflex contraction of the surrounding muscles. This can make relocating a dislocated joint impossible without paralysis. Even more obviously, an uninvited cough or wriggle during brain or eye surgery may be disastrous and a patient is paralysed prophylactically.
Paralysis is achieved using two different types of agent: depolarizing and non-depolarizing neuromuscular blockers. The former is an analogue of ACh itself and activates the nicotinic ACh receptor at the NMJ. Unlike ACh however, the associated cation channel is maintained in its open state so that after the initial depolarization and twitch, the muscle fibre is held in its refractory, relaxed state. These agents have the advantage that they act quickly and then wear off in a few minutes, ideal for a very brief intervention such as endotracheal intubation. However, they have a physiologically interesting danger. ACh receptors are usually tightly clustered on the muscle fibre surface at the motor endplate. But diseases which result in ‘denervation’ – a reduction in the number or activity of the motor nerves reaching the muscle (such as spinal trauma or Guillain-Barré syndrome) – trigger a compensatory increase in the number of functional receptors which appear all over the entire muscle fibre surface. Administration of a depolarizing agent in this context can liberate enough intracellular potassium to cause hyperkalaemia, cardiac arrest and death.
Non-depolarizing agents are analogues of the poison curare which is derived from tree frogs and used to hunt in certain parts of the world. They act simply by sitting on the ACh receptor at the motor endplate and preventing the ACh liberated from the presynaptic membrane from reaching the receptor, activating the cation channel and depolarizing the fibre. They have no intrinsic activity of their own. It is possible to reverse the action of non-depolarizing neuromuscular blocking agents by administering an acetylcholinesterase inhibitor, which allows enough ACh to accumulate in the NMJ to overcome the receptor blockade.
Central excitatory synapses
Once the principles of operation of the NMJ are understood, the operation of the synapses by which one neuron can excite another presents little extra difficulty. The general sequence of events is identical: vesicles containing neurotransmitter are released from the presynaptic terminal by an influx of calcium triggered by depolarization; these vesicles were manufactured in the Golgi apparatus of the cell body and transported to the terminal using microtubules (though one difference is that the incorporation of small transmitter molecules into the vesicles may occur in the terminal rather than in the cell body). When released, the transmitter is recognized by receptors located in the postsynaptic membrane, that in turn alter the permeability of ion channels, leading to the generation of currents and voltages, and other effects. This whole sequence of events is summarized in the diagram of the schematic neuron on p. 47.
The most frequently studied synapse of this kind is one forming part of the monosynaptic reflex arc in the spinal cord that generates the tendon jerk response. This reflex pathway, whose functions will be discussed in Chapter 10, consists of exactly two neurons: a primary (Ia) afferent fibre carrying impulses from stretch receptors in a muscle synapses excitatorily with a motor neuron in the ventral horn of the spinal cord, whose axon returns to innervate the same muscle from which the afferent fibre came. Tapping the tendon of the muscle causes a brief stretch of the sensory ending, firing the Ia fibre, which then excites the motor neuron and causes a reflex twitch of the muscle – the familiar knee-jerk response, if we use the patellar tendon.
The advantage of this reflex pathway from the experimenter’s point of view is that the afferent fibres are readily accessible in the dorsal root for controlled stimulation, while the postsynaptic cell bodies are large enough to be punctured easily by a microelectrode. If we apply a single brief shock to the Ia fibres while recording from the motor neuron, we find that the postsynaptic response consists of a small depolarization rather similar in shape to the EPP, called the excitatory postsynaptic potential (EPSP); if it is large enough, it may trigger off an impulse in the motor neuron. By measuring the reversal potential in exactly the same way as in the case of the NMJ, it is possible to show that the EPSP is the result of a transient increase in permeability to sodium and potassium ions. Just as in the case of the endplate, although this increase lasts only about as long as the action potential, the EPSP itself is relatively prolonged because of the long time constant of the cell membrane. One may therefore assume that the arrival of an impulse releases some transmitter substance from the vesicles visible in the presynaptic endings and that this substance diffuses across the synaptic cleft and causes the opening of short-circuiting channels. Could this transmitter also be ACh?
It would be nice if we could simply extract the vesicles from the ending and see what was in them: but this is technically difficult. However, there are histological stains (and more recently and usefully, immunohistochemical stains) that are selective for particular transmitters or their metabolic precursors, or for enzymes that are associated with them, which can help to identify transmitter substances. In the case of ACh, which is widely found as a transmitter within the brain as well as at the NMJ, one may stain for the enzyme cholinesterase, whose presence suggests strongly the use of ACh itself. But the mere presence of a possible transmitter in the presynaptic endings is not sufficient evidence by itself that it is actually being used as a transmitter. There are a number of further criteria that have to be met. We need to confirm, for example, that application of the supposed transmitter actually causes the same effects as the real transmitter. It is not enough simply to note that both are excitatory: they must both open the same channels, and so have the same reversal potential. Ideally it should also do so in plausibly small concentrations, but this is a difficult criterion to meet because the postsynaptic membrane is very much less accessible from outside than it is to transmitter released in the proper way from the terminal. We must also demonstrate that the real and supposed transmitters have the same pharmacology: that they are blocked by the same pharmacological agents, and that substances that inhibit the inactivating enzyme for one, and hence prolong its action, do so for the other. Ideally, one should also be able to show that afferent action potentials really do release the supposed transmitter, but this also is often technically extremely difficult to establish adequately.
In fact it is only at a relatively small proportion of the synapses in the central nervous system that we are absolutely certain of the identity of the transmitter in the sense that all these criteria have been met. But if one is satisfied with circumstantial evidence, then one can generate quite long lists of putative transmitters (Box 3.1) and create maps indicating their distribution throughout the brain. It would be nice to think that such maps would reveal some deep pattern of meaning as regards which transmitter does what, but disappointingly this is not really so. Knowing what the transmitter is at a particular synapse does not generally help us to understand what the synapse does, not least because that is a function of what the receptor site is linked to: the same transmitter may do several quite different things that bear no obvious functional relation to each other.
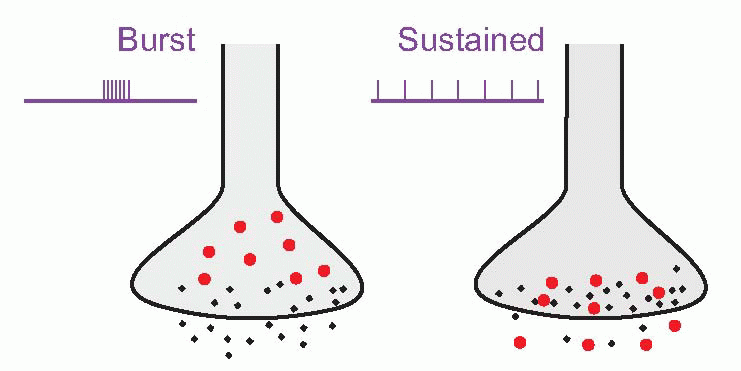
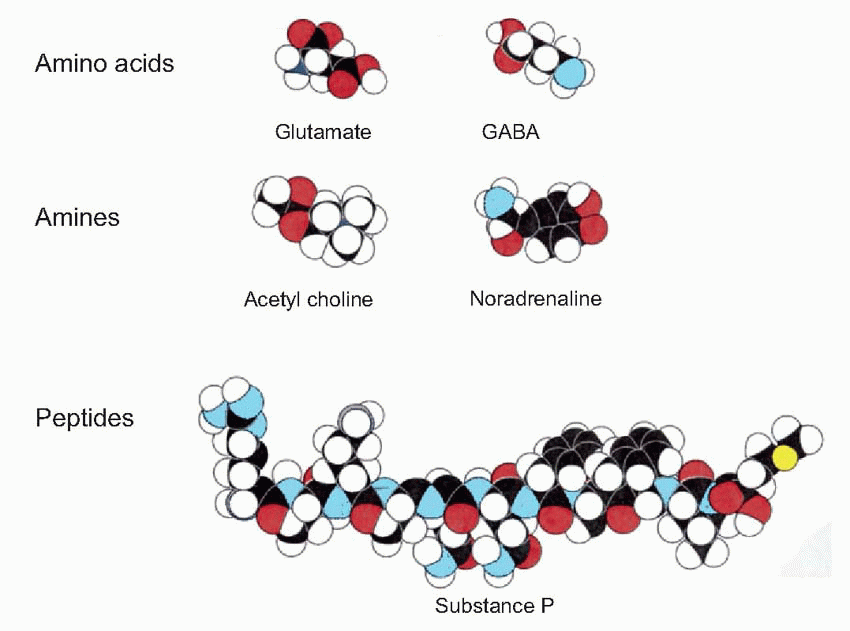
In addition, some synaptic terminals release more than one type of molecule, often a conventional transmitter in conjunction with one or more small peptides such as substance P, vasoactive intestinal peptide (VIP), cholecystokinin (CCK), etc., called co-transmitters, often acting as neuromodulators. Examples of co-transmitters are VIP and substance P with ACh, and enkephalin and neuropeptide Y with noradrenaline. Because the peptides are often packaged in larger vesicles (shown in red, below), further from the synaptic junction than the conventional transmitter (black), it has been suggested that different patterns of afferent stimulation could result in different proportions of the two kinds of transmitter being released; sustained activity would favour the peptide, bursts of activity the conventional transmitter. Some evidence that this actually happens has been found in peripheral autonomic synapses; if it is true, the functional implications are profound, since it would provide a mechanism potentially capable of discriminating more subtle aspects of the action potential code than simply the mean frequency of firing. We shall see later (p. 59) that many neurons in the CNS have mechanisms by which they can generate more complex patterns of firing such as periodic bursts. In addition, there is increasing evidence that neuropeptides frequently have different kinds of effects from ‘conventional’ transmitters. As we shall see later, they often appear to modify the action of transmitters, rather than initiating action themselves, and sometimes they are released in a more diffuse way than is typical for conventional transmitters, suggesting a function that is almost intermediate between neurotransmitter and hormone. (Many of them are of course hormones in other contexts.) Conventional transmitters – ACh and 5-HT for instance – may also act as diffusely acting neuromodulators.
Box 3.1 Examples of types of neurotransmitters and modulators
Neuropeptides (size in brackets):
β-endorphin* (31)
Enkephalins* (5)
Oxytocin* (9)
Glucagon* (29)
Somatostatin* (14)
Substance P (11)
VIP (vasoactive intestinal polypeptide)* (28) (and many others)
Amino acids:
Aspartate
Glutamate
GABA
Glycine
Monoamines:
Dopamine
Noradrenaline*
5-HT (5-hydroxytryptamine, serotonin)*
Other:
ACh
Nitric oxide?
Carbon monoxide?
ATP?
*Also used as a hormone.
Table 3.2 Some varieties of receptors for neurotransmitters


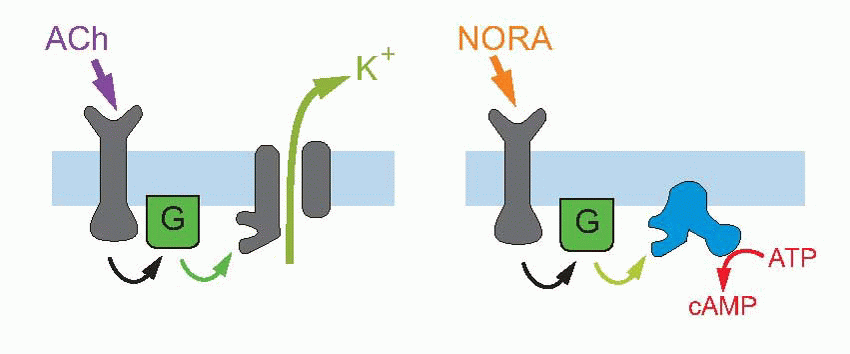
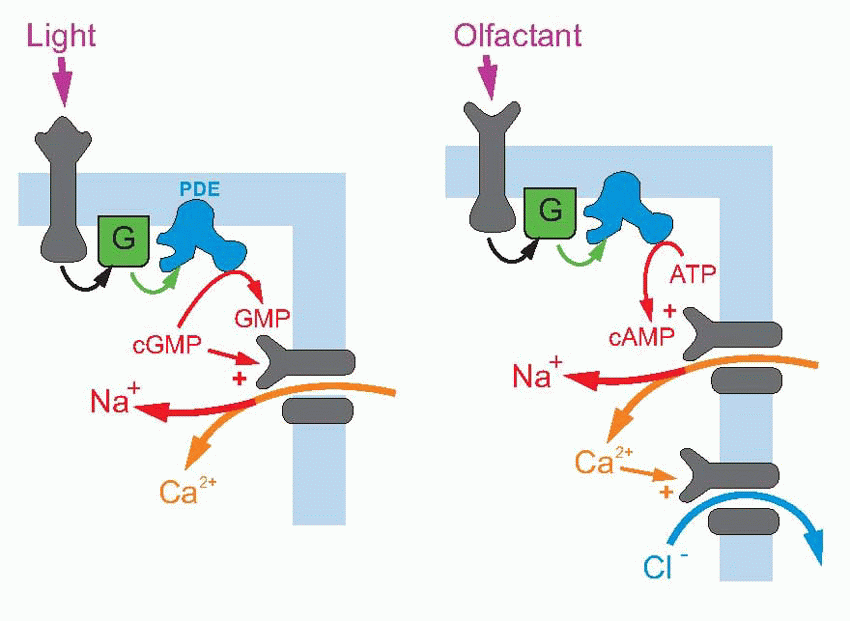
 where intracellular calcium modifies the sensitivity and time-course of the cGMP changes, thus effectively altering the receptor’s sensitivity to light. An obvious example of intracellular effects is the β-adrenergic response to noradrenaline, where cAMP production is again coupled via a G-protein to the receptor itself, the cAMP then having metabolic effects within the cell.
where intracellular calcium modifies the sensitivity and time-course of the cGMP changes, thus effectively altering the receptor’s sensitivity to light. An obvious example of intracellular effects is the β-adrenergic response to noradrenaline, where cAMP production is again coupled via a G-protein to the receptor itself, the cAMP then having metabolic effects within the cell.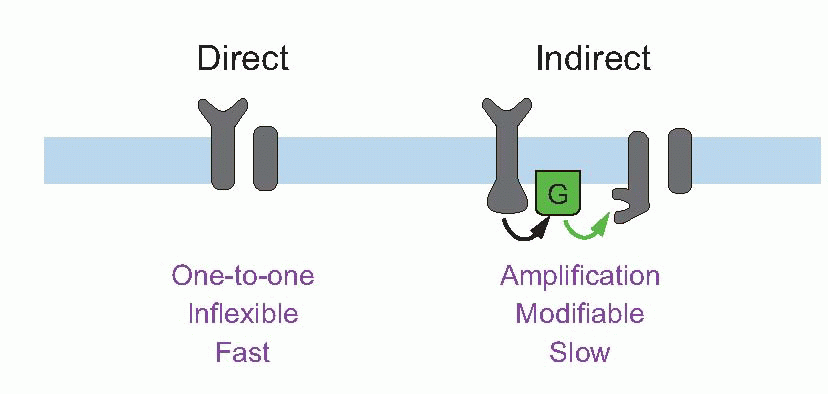
 The fact that the current hardly lasts longer than the afferent impulse is largely due to the presence at the ending of high concentrations of the enzyme cholinesterase that mops up the ACh almost as soon as it arrives. If this enzyme is blocked by an anticholinesterase such as eserine, one finds that the current flow, and hence the EPP, is enormously prolonged, leading to a depolarization block of the muscle fibre. Patch-clamping demonstrates that the channels operate in an all-or-nothing manner, being either fully open or fully closed, as can be seen in the recording from rat NMJ below – which also shows the increase in the probability of opening with increasing calcium concentration. In addition, this technique can be used to demonstrate that two ACh molecules are required to trigger the opening of one channel.
The fact that the current hardly lasts longer than the afferent impulse is largely due to the presence at the ending of high concentrations of the enzyme cholinesterase that mops up the ACh almost as soon as it arrives. If this enzyme is blocked by an anticholinesterase such as eserine, one finds that the current flow, and hence the EPP, is enormously prolonged, leading to a depolarization block of the muscle fibre. Patch-clamping demonstrates that the channels operate in an all-or-nothing manner, being either fully open or fully closed, as can be seen in the recording from rat NMJ below – which also shows the increase in the probability of opening with increasing calcium concentration. In addition, this technique can be used to demonstrate that two ACh molecules are required to trigger the opening of one channel.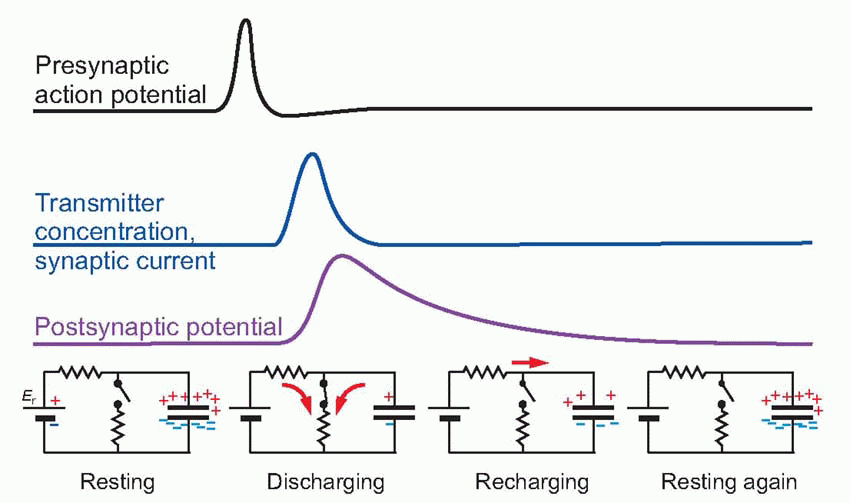
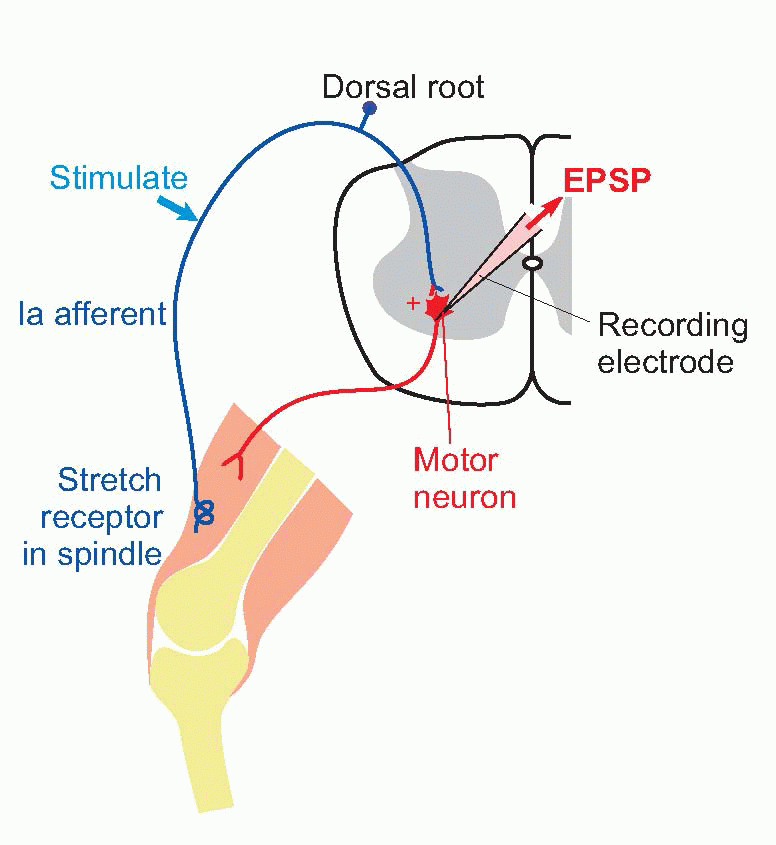
 If we apply a single brief shock to the Ia fibres while recording from the motor neuron, we find that the postsynaptic response consists of a small depolarization rather similar in shape to the EPP, called the excitatory postsynaptic potential (EPSP); if it is large enough, it may trigger off an impulse in the motor neuron. By measuring the reversal potential in exactly the same way as in the case of the NMJ, it is possible to show that the EPSP is the result of a transient increase in permeability to sodium and potassium ions. Just as in the case of the endplate, although this increase lasts only about as long as the action potential, the EPSP itself is relatively prolonged because of the long time constant of the cell membrane. One may therefore assume that the arrival of an impulse releases some transmitter substance from the vesicles visible in the presynaptic endings and that this substance diffuses across the synaptic cleft and causes the opening of short-circuiting channels. Could this transmitter also be ACh?
If we apply a single brief shock to the Ia fibres while recording from the motor neuron, we find that the postsynaptic response consists of a small depolarization rather similar in shape to the EPP, called the excitatory postsynaptic potential (EPSP); if it is large enough, it may trigger off an impulse in the motor neuron. By measuring the reversal potential in exactly the same way as in the case of the NMJ, it is possible to show that the EPSP is the result of a transient increase in permeability to sodium and potassium ions. Just as in the case of the endplate, although this increase lasts only about as long as the action potential, the EPSP itself is relatively prolonged because of the long time constant of the cell membrane. One may therefore assume that the arrival of an impulse releases some transmitter substance from the vesicles visible in the presynaptic endings and that this substance diffuses across the synaptic cleft and causes the opening of short-circuiting channels. Could this transmitter also be ACh?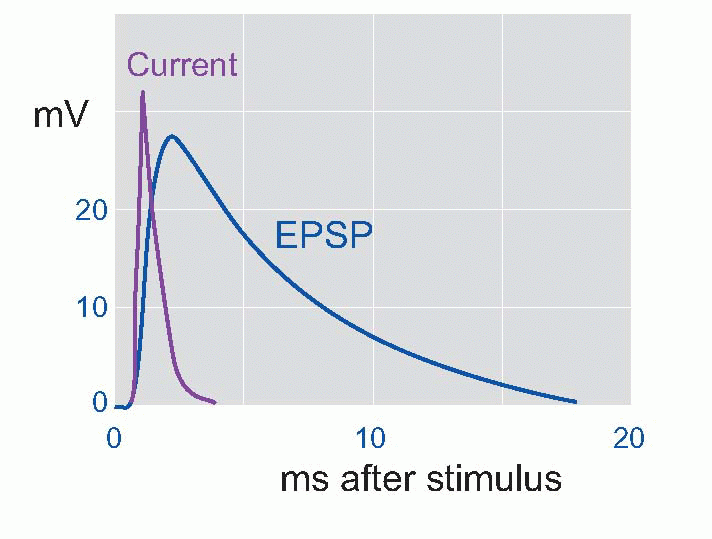
 It would be nice to think that such maps would reveal some deep pattern of meaning as regards which transmitter does what, but disappointingly this is not really so. Knowing what the transmitter is at a particular synapse does not generally help us to understand what the synapse does, not least because that is a function of what the receptor site is linked to: the same transmitter may do several quite different things that bear no obvious functional relation to each other.
It would be nice to think that such maps would reveal some deep pattern of meaning as regards which transmitter does what, but disappointingly this is not really so. Knowing what the transmitter is at a particular synapse does not generally help us to understand what the synapse does, not least because that is a function of what the receptor site is linked to: the same transmitter may do several quite different things that bear no obvious functional relation to each other.