Chapter 21 Control of Eye Movements
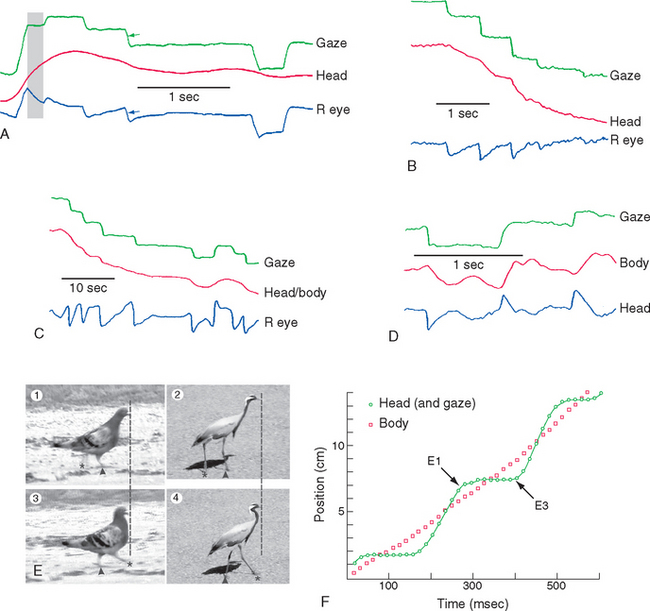
Photoreceptors are sensitive but slow, and all animals with image-forming eyes have mechanisms to prevent images of interest from moving across the retina too quickly to be analyzed. (Using the camera analogy from Chapter 17, our eyes behave in many respects like cameras with a shutter speed of about 1/10 of a second.) A widespread strategy is to use some combination of eye movements and head or body movements to keep the direction of gaze constant, except for brief periods during which gaze shifts (Fig. 21-1). Additional complications are added by having a fovea, which results in good spatial acuity for only a small area of central vision (see Fig. 17-18). Finally, effective binocular vision requires precise alignment of the two eyes. All this necessitates a highly accurate system for controlling eye position and movement.
Six Extraocular Muscles Move the Eye in the Orbit
Six small extraocular muscles (Fig. 21-2) rotate each eye in its orbit, like a ball in a socket. Four rectus (“straight”) muscles (medial, lateral, superior, and inferior) originate from a common tendinous ring (the annulus of Zinn) in the back of the orbit and insert anteriorly in the sclera, 5 to 8 mm from the limbus. Two oblique muscles, as the name implies, pass obliquely over the surface of the eye and insert in the sclera of its posterior half. The superior oblique originates near the common tendinous ring, from the sphenoid bone at the back of the orbit, but it takes an unusual course in reaching the eye. Near the front of the orbit, the superior oblique tendon passes through a fibrous loop (the trochlea*) attached to the frontal bone, turns laterally and posteriorly, and inserts in the sclera of the posterior half of the eye. The inferior oblique originates anteriorly and medially from the floor of the orbit, passes across the inferior surface of the eye, and inserts posteriorly.
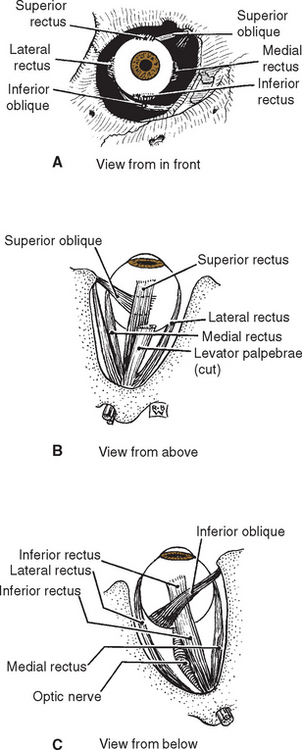
Figure 21-2 Anterior (A), superior (B), and inferior (C) views of the extraocular muscles of the right eye.
(Modified from von Noorden GK: Atlas of strabismus, ed 4, St. Louis, 1983, Mosby.)
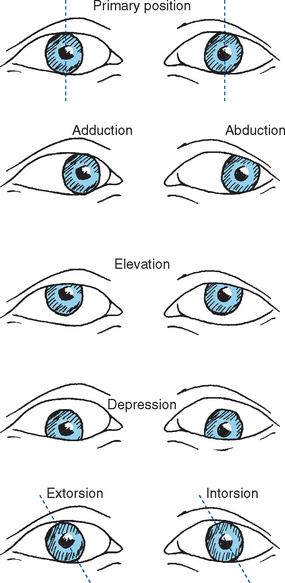
Different patterns of contraction of the six extrao-cular muscles rotate the eye horizontally (adduction, abduction), vertically (elevation, depression), or around its anterior-posterior axis (extorsion, intorsion) (Fig. 21-3).
The Medial and Lateral Recti Adduct and Abduct the Eye
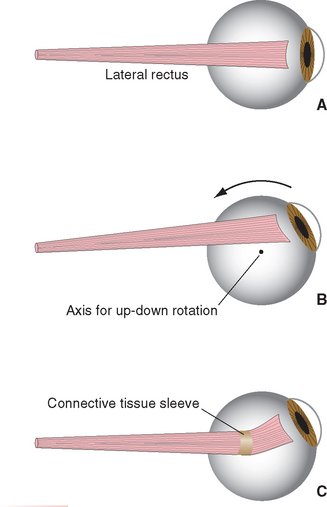
The medial and lateral recti are situated in a horizontal plane, so contractions of these muscles rotate the eye around a vertical axis. This makes their actions straightforward (Fig. 21-4)—the medial rectus adducts and the lateral rectus abducts the eye. Both muscles pass through sleeves of connective tissue (referred to as pulleys) as they travel from the common tendinous ring toward the eye, emerging a little posterior to their insertion points on the sclera. The pulleys, in turn, have connective tissue attachments to the walls of the orbit, so the positions of the muscles are constrained within the orbit as the eye moves. As a result, adduction and abduction continue to be the principal effects of the medial and lateral recti, even if the eye starts out in an elevated or depressed position (Fig. 21-5). The superior and inferior recti and the inferior oblique have similar pulleys (so in this sense, the superior oblique is not as different from the other extraocular muscles as it seems to be). The pulleys form important parts of an elaborate mechanical suspension system that helps control rotations of the eye; half the fibers of the recti and inferior oblique actually attach to the pulleys rather than the sclera, adjusting pulley positions as the eye moves.
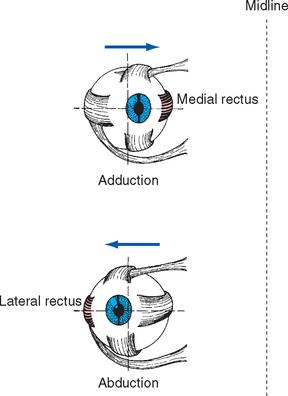
Figure 21-4 Actions of the medial and lateral recti of the right eye.
(Modified from Moses RA, editor: Adler’s physiology of the eye, ed 5, St. Louis, 1970, Mosby.)
The Superior and Inferior Recti and the Obliques Have More Complex Actions
The actions of the superior and inferior recti and the obliques are not quite so straightforward, because the anatomical axis of the orbit is deviated about 23 degrees laterally from the visual axis of the eye (Fig. 21-6). The result, as indicated for the superior rectus and superior oblique in Figure 21-7 but equally true for the inferior rectus and inferior oblique, is that when one of these muscles contracts, it looks as though the eye should rotate around an axis that is neither vertical nor transverse nor anterior-posterior. Consequently, each of these four muscles has one principal action and weaker additional actions (Fig. 21-8). However, the pulley system constrains their directions of pull, and these secondary actions come into play only in eccentric gaze. Starting from straight-ahead gaze, the superior and inferior recti largely take care of elevation and depression, respectively; similarly, the superior and inferior obliques cause intorsion and extorsion.
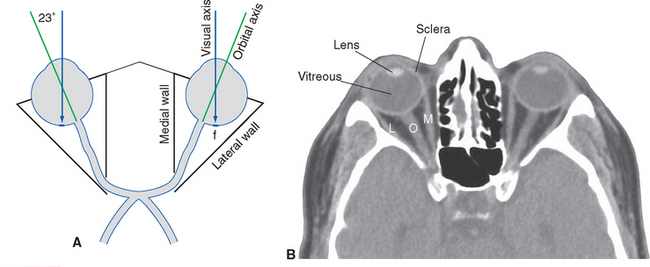
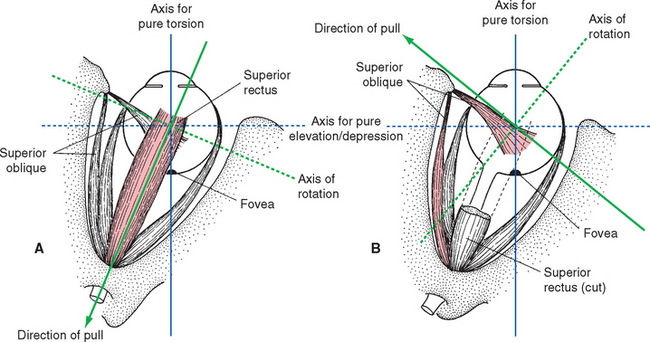
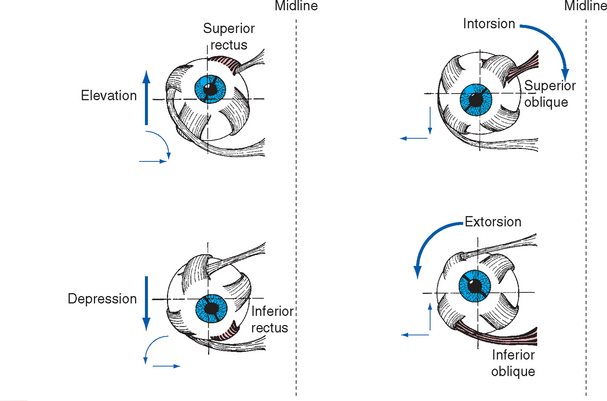
Figure 21-8 Actions of the right superior and inferior recti and obliques. Each has a primary action (large arrows) and two secondary actions (small arrows) when starting from the primary position. The actions are determined both by the direction of insertion of a muscle’s tendon and by the position of the insertion. For example, the superior rectus and superior oblique both insert on the superior surface of the globe. However, the superior rectus inserts anteriorly and pulls posteriorly, so it elevates the eye. In contrast, the superior oblique inserts posteriorly and pulls in part anteriorly (Fig. 21-7), so part of its action is depression. The extent of the secondary actions of the superior and inferior recti is somewhat exaggerated in this figure; connective tissue pulleys constrain their actions to nearly pure elevation and depression, respectively, in straight-ahead gaze.
(Modified from Moses RA, editor: Adler’s physiology of the eye, ed 5, St. Louis, 1970, Mosby.)
To keep things reasonably simple in this chapter, torsional movements are not discussed,* and adduction, abduction, elevation, and depression are treated as the result of contraction of just one of the rectus muscles. However, eye movements actually involve coordinated changes in the activity of all six extraocular muscles (Table 21-1), some contracting and some relaxing in response to changing levels of input from the oculomotor, trochlear, and abducens nuclei (see Fig. 12-4 to 12-6). For example, adduction involves simultaneous contraction of the medial, superior, and inferior recti and relaxation of the lateral rectus and the superior and inferior obliques; the intorsion and extorsion of the superior and inferior recti cancel each other.
Table 21-1 Extraocular Muscles Contributing to Movements Around Different Axes
| Movement | Principal Muscle | Other Muscles Contributing |
|---|---|---|
| Adduction | Medial rectus | Superior rectus |
| Inferior rectus | ||
| Abduction | Lateral rectus | Superior oblique |
| Inferior oblique | ||
| Elevation | Superior rectus | Inferior oblique |
| Depression | Inferior rectus | Superior oblique |
| Extorsion | Inferior oblique | Inferior rectus |
| Intorsion | Superior oblique | Superior rectus |
There Are Fast and Slow Conjugate Eye Movements
Conjugate movements serve one of two general purposes—to move an object’s image onto the fovea or to keep it there (Fig. 21-1). We use fast, steplike eye movements called saccades (from a French word meaning “a pull on the reins”) to redirect gaze so a different image falls on the fovea. The same neural machinery comes into play whenever these brief, rapid movements are called for—to move our eyes voluntarily in any given direction, to glance over at something in the periphery, and in the fast phase of nystagmus. We use multiple kinds of smooth, slower eye movements to keep an image on the fovea, corresponding to the fact that an image can move on the retina if we move or if the object moves. The vestibuloocular reflex (VOR) (see Figs. 14-31 and 14-32) compensates for head movements, supplemented in this function by optokinetic movements (see Fig. 14-33A); for large head movements, these smooth eye movements become the slow phase of nystagmus. Smooth pursuit movements are used to track a visual stimulus that is itself moving. Without special training, we are unable to move our eyes smoothly unless an image starts to leave the fovea (Box 21-1).
It generally comes as a surprise that, with the head stationary, we cannot voluntarily move our eyes smoothly unless we are tracking a slowly moving object (Fig. 21-9A). This is easily demonstrated, however. Watch someone’s eyes as he or she tries to move them slowly and smoothly while there is nothing to track, or concentrate on your own eyes while you try to do the same thing. In either case the result will be the same: a series of rapid, jerky movements (saccades; Fig. 21-9B). Some individuals can learn to have voluntary control of smooth tracking movements, but under normal circumstances and for most people, this is impossible. Interestingly, real moving stimuli in other sensory modalities provide, to varying degrees, an adequate stimulus for some smooth movements. For example, many people can use smooth eye movements to track their own index fingers moving back and forth in the dark (Fig. 21-9C), even though they are unable to use such eye movements to track imaginary moving fingers. Similarly, smooth eye movements can be used to track a moving sound source in the dark (although less successfully than when using a somatosensory stimulus).
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


