Chapter 12 Cranial Nerves and Their Nuclei
Cranial Nerve Nuclei Have a Generally Predictable Arrangement
Spinal nerves contain sensory and motor fibers. Some of each kind are related to visceral structures and some to somatic structures. A given axon entering or leaving the spinal cord can therefore be placed in one of the following four categories:
By and large the cell bodies on which spinal afferents synapse and the cell bodies of spinal efferent fibers are located in portions of the spinal gray matter predictable from its embryological development (Fig. 12-1A). The sulcus limitans separates the alar plate (which develops into the posterior horn) from the basal plate (which develops into the anterior horn). Within both the alar and the basal plates, cells concerned with visceral function tend to be located nearer the sulcus limitans. This is shown most clearly in the adult central nervous system (CNS) by the location of the cell bodies of visceral motor neurons in the intermediolateral cell column. So for each of the four spinal axon categories there is a corresponding column of cells in the spinal gray matter. The somatic sensory and motor columns extend the length of the cord; the visceral sensory and motor columns are found at spinal levels T1 to L2-L3 and S2 to S4.
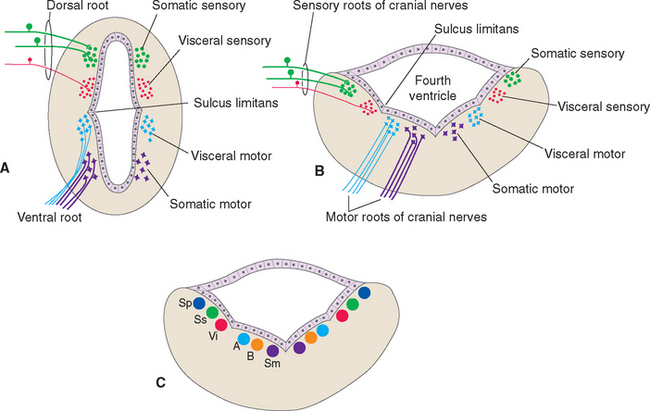
The Sulcus Limitans Intervenes between Motor and Sensory Nuclei of Cranial Nerves
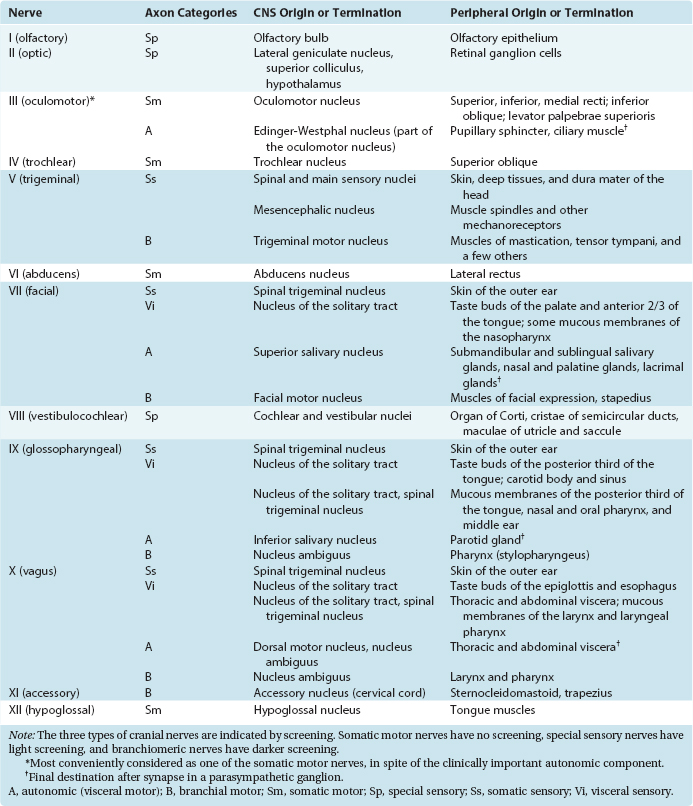
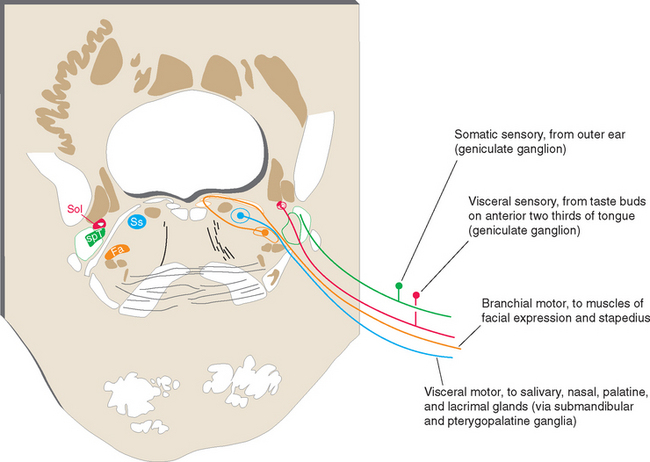
Axons from all four categories found in spinal nerves are also found in various cranial nerves, where they take care of the same functions for the head. However, some cranial nerves contain axons from additional categories, reflecting specialized structures and functions associated with the head. Thus there are special sensory fibers that, in the case of the cranial nerves attached to the brainstem, are related to the special senses of hearing and equilibrium. * In addition, motor axons in certain cranial nerves innervate striated muscles with a special embryological origin, referred to as branchiomeric muscles. Structures that develop into the gill arches (or branchial arches) in fish develop instead into various structures in and near the head and neck in humans and other mammals. Branchiomeric muscles (notably the muscles of the larynx, pharynx, jaw, and face) are associated with these branchial arch structures. Functionally and histologically, branchiomeric muscles are identical to ordinary skeletal muscle, but the motor neurons for branchiomeric muscles have a distinctive location in the brainstem, different from that of ordinary somatic motor neurons. In recognition of their special development and location, they are classified as a separate category, here called branchial motor neurons.* Hence there are six different categories of nerve fibers in the cranial nerves attached to the brainstem (Table 12-1).
Table 12-1 Categories of Nerve Fibers in Cranial Nerves of the Brainstem
| Structures Innervated | Cranial Nerves* | |
|---|---|---|
| Sensory | ||
| Somatic | Skin, muscles, joints of the head | V |
| Visceral | Cranial, thoracic, abdominal viscera | X |
| Taste buds | VII, IX | |
| Special | Inner ear | VIII |
| Motor† | ||
| Somatic | Extraocular muscles | III, IV, VI |
| Tongue muscles | XII | |
| Visceral | Parasympathetic ganglia for cranial, thoracic, and abdominal viscera | X |
| Branchial | Jaw muscles | V |
| Facial muscles | VII | |
| Laryngeal and pharyngeal muscles | X | |
| Middle ear muscles | V, VII | |
| Sternocleidomastoid, trapezius | XI | |
* Principal cranial nerves only. Smaller contributions that may nevertheless be clinically important (e.g., parasympathetics for the pupil in cranial nerve III) are not indicated here but are included in Table 12-2.
† Does not include the efferents in cranial nerve VIII (described in Chapter 14) that innervate the receptor cells of the inner ear, which do not fit comfortably into any of these categories.
As in the case of the spinal cord, the locations of the cell bodies where cranial nerve afferents terminate or cranial nerve efferents originate can be predicted, to some extent, from the embryology of the brainstem. The walls of the neural tube spread apart in the medulla and pons to form the floor of the fourth ventricle (see Fig. 2-10). The sulcus limitans runs longitudinally along the floor of the adult ventricle (see Fig. 11-3A), still separating sensory alar plate derivatives (now lateral) from motor basal plate derivatives (now medial) (Fig. 12-1B). As in the case of the spinal cord, cells concerned with visceral functions are usually located nearer the sulcus limitans.
Ideally the cell columns subserving the special components of the cranial nerves would be located adjacent to those for the corresponding general components, as indicated in Figure 12-1C. The actual arrangement in the adult brainstem is not quite as simple as in this idealized diagram, for two principal reasons. First, the cell columns of the brainstem are not continuous like those of the spinal cord; rather, they are interrupted and form a series of nuclei located at longitudinal levels roughly corresponding to the attachment points of the cranial nerves. As a result, all components are seldom present in a given transverse plane (Fig. 12-2; see also Fig. 15-2). Second, in a few instances, portions of a cell column migrate away from their expected locations (Fig. 12-3). For example, most branchial motor neurons are located in the ventrolateral part of the tegmentum rather than in the floor of the ventricle adjacent to other efferent neurons. The actual locations of cranial nerve nuclei in the rostral medulla are shown in Figure 12-3; also indicated are the functional types of fibers in each of the cranial nerves of the brainstem. (This is only meant to be a convenient summary; not all cranial nerves project to or originate from the rostral medulla.)
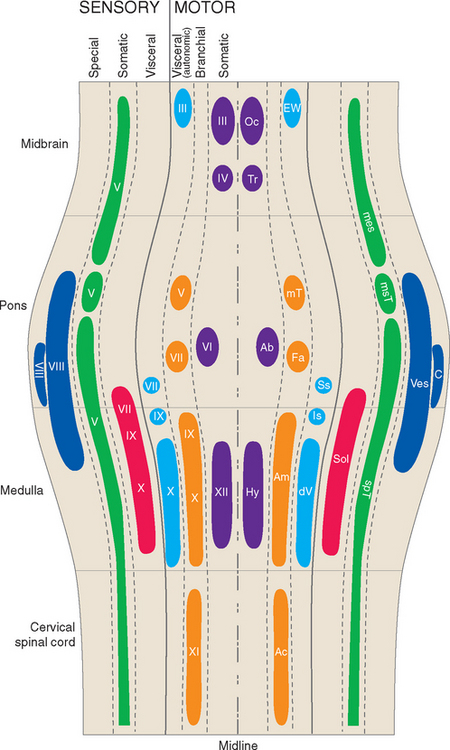
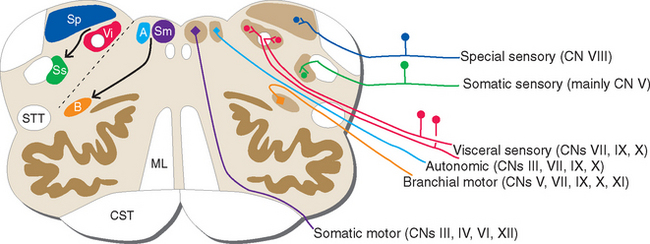
It can be seen from Figure 12-3 that no cranial nerve contains axons from all six categories. If the compositions of all the nerves are tabulated (as in Table 12-2), it becomes apparent that there are three types of cranial nerves. Some nerves (III, IV, VI, and XII) contain motor axons for ordinary skeletal muscle and little or nothing else, so they may be referred to as somatic motor nerves. Others (I, II, and VIII) contain special sensory fibers and little or nothing else. The remaining nerves (V, VII, IX, X, and XI) are somewhat more complex and typically contain several components; all innervate branchial arch musculature, so they are called branchiomeric nerves.
A presentation of the cranial nerves involves too much material for one comfortable sitting, so it is spread out over several chapters. The remainder of this chapter is divided into two more or less distinct sections, discussing first the somatic motor nerves and then most components of the branchiomeric nerves. Chapter 13 discusses the chemical senses of taste and smell subserved by some brainstem cranial nerves and the olfactory nerve. Chapter 14 deals with the eighth nerve, the special sensory nerve subserving hearing and equilibrium. (The remaining special sensory nerve, the optic nerve, is an outgrowth of the diencephalon and is really a tract of the CNS. It is considered separately in Chapter 17.) Finally, as mentioned previously, Chapter 15 contains a series of brainstem sections with labels and summary descriptions indicating the locations and contents of cranial nerve nuclei and other important brainstem structures.
Cranial Nerves III, IV, VI, and XII Contain Somatic Motor Fibers
The somatic motor nerves are the simplest of the cranial nerves because each contains fibers of only one category (except for cranial nerve III, which has a small but important complement of preganglionic parasympathetic fibers). * The nuclei of origin of all these nerves are located adjacent to the midline near the aqueduct or the floor of the fourth ventricle, as would be expected from their embryological origins.
The Oculomotor Nerve (III) Innervates Four of the Six Extraocular Muscles
Cranial nerve III supplies the levator palpebrae superioris (the principal elevator of the eyelid) and all the internal and external muscles of the ipsilateral eye except the lateral rectus, superior oblique, and pupillary dilator. The fibers originate in the wedge-shaped oculomotor nucleus, which is located at the ventral edge of the periaqueductal gray in the rostral midbrain (Fig. 12-4). They then proceed ventrally and arch through the midbrain tegmentum in several separate bundles that join to form the nerve just as they emerge into the interpeduncular fossa.
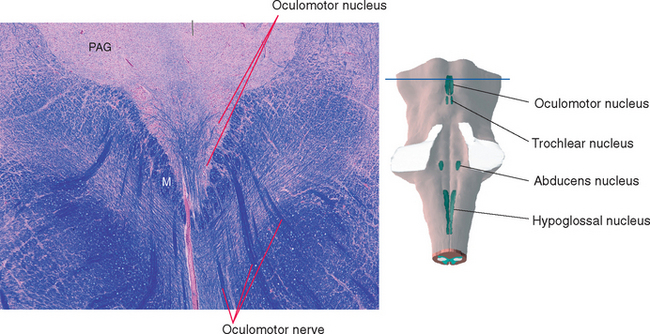
Figure 12-4 Section through the rostral midbrain showing the oculomotor nucleus. This figure is an enlargement of part of the section shown in Figure 11-17. The oculomotor nucleus is actually a tight cluster of subnuclei, each of which innervates a different muscle. M, medial longitudinal fasciculus; PAG, periaqueductal gray.
The partly crossed–partly uncrossed nature of the oculomotor nerve is a curious fact but one of limited clinical significance. This is because the oculomotor nuclei of the two sides are so close to each other that a central lesion in this vicinity is likely to damage both nuclei. However, once a given oculomotor nerve emerges from the brainstem, it supplies only ipsilateral muscles, so a lesion of the third nerve, or of fibers curving through the midbrain tegmentum on their way to the third nerve, affects only one eye. Therefore the dissociated findings of paralysis of the superior rectus on one side and of other extraocular muscles on the opposite side are rarely encountered (although there are occasional cases of unilateral nuclear damage in which these deficits are found).
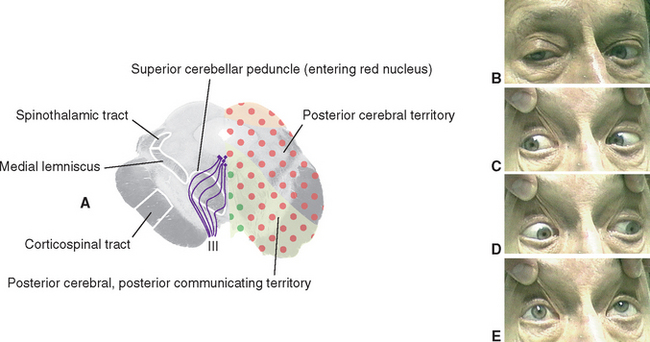
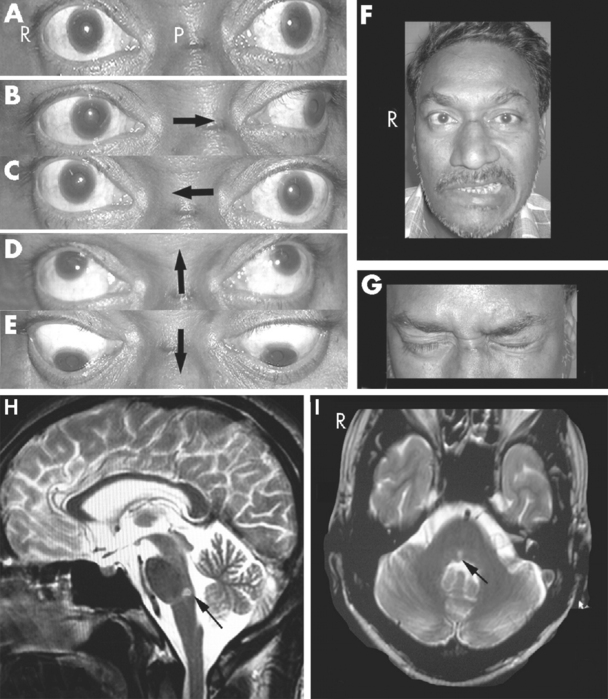
Damage to one oculomotor nerve causes a series of deficits (see Fig. 12-31B to E). The eye ipsilateral to the lesion deviates laterally because the medial rectus is now paralyzed and the lateral rectus is unopposed. This is called lateral strabismus, indicating that the eyes are misaligned because one of them deviates laterally from midposition. As a result, the patient complains of diplopia (double vision) and is unable to move the affected eye medially; vertical movements are also impaired because of paralysis of the superior and inferior recti and the inferior oblique. The ipsilateral levator palpebrae superioris is paralyzed, so ptosis occurs. In addition, the pupillary sphincter and ciliary muscle are nonfunctional. The pupil on the affected side is dilated (mydriasis) as a result of the now-unopposed pupillary dilator, and it does not constrict in response to light;* the lens cannot be focused for near vision.
The Trochlear Nerve (IV) Innervates the Superior Oblique
Cranial nerve IV, the trochlear nerve, supplies the superior oblique and is named for the sling of connective tissue (the trochlea—Latin for “pulley”) through which the tendon of the superior oblique passes (see Fig. 21-2). Its cell bodies of origin are located in the contralateral trochlear nucleus. This is a small nucleus (because it has only one small muscle to supply) located at the level of the inferior colliculus, where it indents the medial longitudinal fasciculus (MLF) (Fig. 12-5). Fibers leaving the nucleus turn caudally in the periaqueductal gray, then arch dorsally to decussate and leave the brainstem at the pons-midbrain junction. The trochlear nerve is thus unique in two respects: it is the only cranial nerve attached to the dorsal surface of the brainstem and the only one to originate entirely from a contralateral nucleus. *
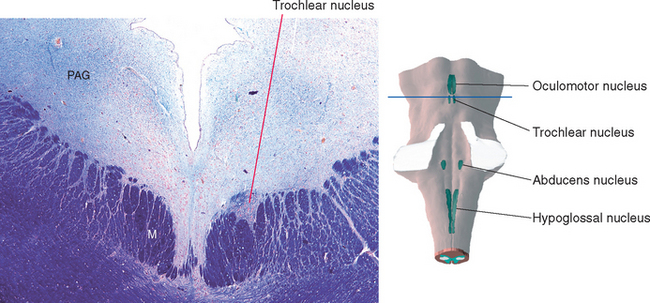
Figure 12-5 Section through the caudal midbrain showing the trochlear nucleus. This figure is an enlargement of part of the section shown in Figure 11-16. M, medial longitudinal fasciculus; PAG, periaqueductal gray.
The Abducens Nerve (VI) Innervates the Lateral Rectus
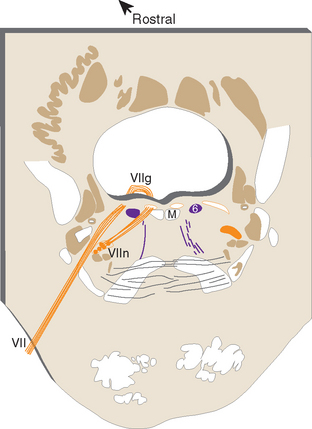
Cranial nerve VI, the abducens nerve, supplies the lateral rectus, which abducts the eye (hence the name of the nerve). The fibers originate from the ipsilateral abducens nucleus, which is located in the caudal pons beneath the floor of the fourth ventricle (Fig. 12-6). Medial to this nucleus are two bundles of fibers. The more medial of the two is the MLF. Between the MLF and the abducens nucleus are motor fibers of the facial nerve, which take an unusual course in leaving the brainstem. They originate in the facial nucleus (see Figs. 12-6 and 12-24), which is located in the ventrolateral part of the pontine tegmentum at about the same level as the abducens nucleus. The facial fibers travel dorsomedially, wrap around the abducens nucleus, and turn back ventrally to exit from the brainstem (Fig. 12-7). The place where these fibers wrap around the abducens nucleus is called the internal genu of the facial nerve. * The abducens nucleus, together with the internal genu, is responsible for the facial colliculus in the floor of the fourth ventricle (see Fig. 11-3A).
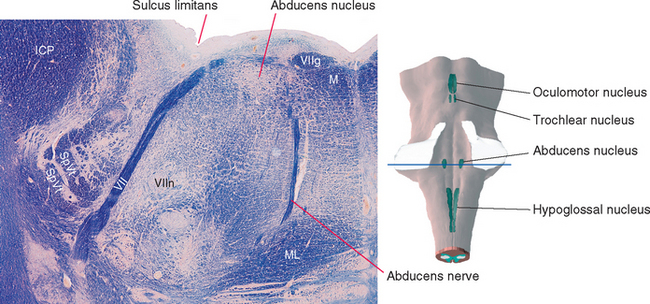
Figure 12-6 Section through the caudal pons showing the abducens nucleus and fibers of the facial nerve cut at different points along their course. The rounded elevation in the floor of the fourth ventricle between the midline and the sulcus limitans is the facial colliculus (see Fig. 11-3A). This figure is an enlargement of part of the section shown in Figure 11-11. ICP, inferior cerebellar peduncle; M, medial longitudinal fasciculus; ML, medial lemniscus; SpVt, spinal trigeminal tract; VII, facial nerve; VIIg, internal genu of the facial nerve; VIIn, facial motor nucleus.
The Abducens Nucleus Also Contains Interneurons That Project to the Contralateral Oculomotor Nucleus
Damage to the abducens nerve causes a medial strabismus (i.e., the affected eye deviates medially) as a result of the action of the now-unopposed medial rectus. The individual may be able to move the affected eye from the adducted position to midposition (but not past it) by relaxing its medial rectus (Fig. 12-8A). Damage to the abducens nucleus causes the same deficit, but with a significant addition: the ipsilateral eye will not abduct past midposition, and the contralateral eye will not adduct past midposition (Fig. 12-8B; see also Fig. 12-10C). This is called lateral gaze paralysis, and it occurs because the abducens nucleus contains not only lateral rectus motor neurons but also an approximately equal number of internuclear neurons with axons that ascend through the MLF.
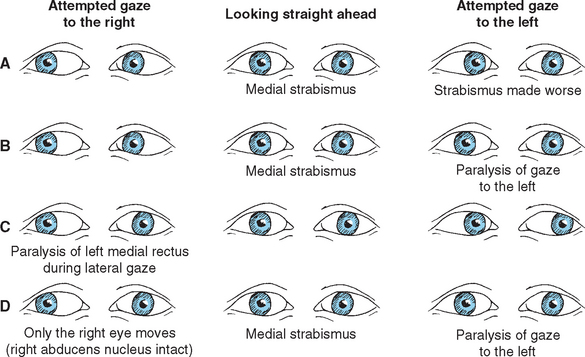
Figure 12-8 Deficits of horizontal gaze after damage to the abducens–MLF system at the sites indicated in Figure 12-9. A, Abducens palsy resulting from damage to the left abducens nerve. B, Lateral gaze paralysis resulting from damage to the left abducens nucleus. C, Internuclear ophthalmoplegia resulting from damage to the left MLF. D, “One-and-a-half” (combination of B and C) resulting from damage to the left abducens nucleus and MLF. In all these situations, convergence, which does not depend on abducens-oculomotor interconnections, is preserved.
The function of the MLF in lateral gaze becomes apparent if you consider that both eyes normally work together: when we look to one side, for example, one lateral rectus and the contralateral medial rectus contract simultaneously. The pathway that interconnects the abducens, trochlear, and oculomotor nuclei to make these sorts of movements possible is the MLF. Vertical movements and the higher centers that direct coordinated eye movements are discussed in Chapter 21, but for purely horizontal movements, the crucial interconnecting fibers are those that arise from the internuclear neurons in the abducens nucleus (Fig. 12-9). These cells send their axons across the midline as they emerge from the abducens nucleus; they then join the contralateral MLF and ascend to the oculomotor nucleus. There they make excitatory synapses on medial rectus motor neurons. Simultaneous firing of abducens motor neurons and internuclear neurons thus results in coordinated lateral gaze.
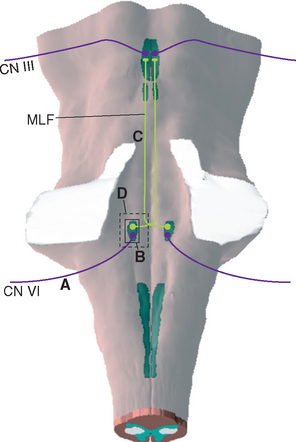
Figure 12-9 Connections between abducens and oculomotor nuclei involved in lateral gaze. Lesions labeled A, B, C, and D would cause the eye movement deficits indicated in Figure 12-8A to D.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


