Hippocampal Anatomy and Hippocampal Sclerosis
Imad Najm
Henri Duvernoy
Stephan Schuele
HIPPOCAMPAL ANATOMY
Because the hippocampal formation is frequently involved in patients with focal epilepsy, a thorough knowledge of this structure and its connections is critical to an understanding of the pathology and clinical semiology of temporal lobe epilepsy caused by hippocampal disease. The first part of this chapter reviews the anatomy of the hippocampal formation, its relation to surrounding structures, and its histology and connections (pathways). The reader is referred to The Human Hippocampus (1) for a comprehensive review. A brief review of the histopathologic changes, mechanisms of epileptogenesis, and neurophysiologic findings in hippocampal sclerosis follows.
The Limbic System
The limbic system is formed by the limbic lobe and associated subcortical structures: amygdala, habenula, mamillary bodies, septal nuclei, and portions of the thalamus, hypothalamus, and midbrain (2, 3, 4, 5). The hippocampus is part of the limbic lobe and is situated on the inferomedial aspect of the hemisphere (Fig. 4.1). The limbic lobe is divided into two gyri (limbic and intralimbic) (6) and is separated from the surrounding cortical structures by the discontinuous limbic fissure, which is composed of cingulate, subparietal, anterior calcarine, collateral, and rhinal sulci. The limbic gyrus consists of the subcallosal, cingulate, and parahippocampal gyri. The latter can be divided into two segments: a narrow posterior segment and a more voluminous anterior segment called the piriform lobe, which comprises the uncus and the entorhinal area. The intralimbic gyrus arches within the limbic gyrus and is composed of two main connected structures: a less developed, narrow, neuronal lamina, which is situated in the anterior (prehippocampal rudiment) and superior surface of the corpus callosum (indusium griseum), and the more developed posterior and ventral hippocampus.
The amygdala is a large nuclear complex in the dorsomedial part of the temporal lobe. It overhangs the hippocampal head along most of its surface and limits the ventral, superior, and medial walls of the temporal horn of the lateral ventricle. The amygdala is continuous with the parahippocampal gyrus caudally and is divided into two major masses of nuclei: the small corticomedial and the well-differentiated basolateral nuclear groups (3). The amygdala is part of the olfactory centers and the limbic system through its connections with both of them.
Hippocampal Anatomy
The hippocampus has the appearance of a sea horse: it bulges into the temporal horn of the lateral ventricle and is arched around the mesencephalon. It is divided into three anatomic segments: the head, which is a transversally oriented segment with prominent digitations (digitationes hippocampi); the body, which is a sagittally oriented segment; and the tail, which is narrow and transversally oriented. The tail disappears beneath the splenium of the corpus callosum posteriorly (Fig. 4.2). The total range of the hippocampus is between 4 and 4.5 cm, the width of the head is between 1.5 and 2 cm, and the width of the body is between 1 and 1.5 cm (7, 8, 9).
The Hippocampal Head
The hippocampal head includes intraventricular and extraventricular (uncal) parts. The intraventricular segment is
anterior within the hippocampus and contains the hippocampal (or internal) digitations.
anterior within the hippocampus and contains the hippocampal (or internal) digitations.
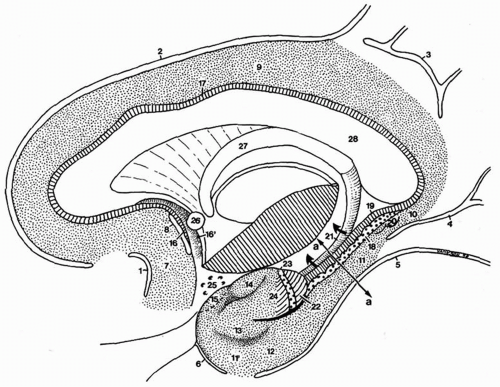 Figure 4.1 Dissection showing a sagittal section of the right hemisphere. The limbic lobe is separated from the isocortex by the limbic fissure and may be divided into two gyri: the limbic and intralimbic gyri. The line (a-a) indicates the plane of the section. Limbic fissure: 1, anterior paraolfactory sulcus (subcallosal sulcus); 2, cingulated sulcus; 3, subparietal sulcus; 4, anterior calcarine sulcus; 5, collateral sulcus; 6, rhinal sulcus. Limbic gyrus: 7, subcallosal gyrus; 8, posterior paraolfactory sulcus; 9, cingulate gyrus; 10, isthmus; 11, parahippocampal gyrus, posterior part; 11′, parahippocampal gyrus, anterior part (piriform lobe). Piriform lobe: 12, entorhinal area; 13, ambient gyrus; 14, semilunar gyrus; 15, prepiriform cortex. Intralimbic gyrus: 16, prehippocampal rudiment; 16′, paraterminal gyrus; 17, indusium griseum. Hippocampus: 18, gyrus dentatus; 19, cornu ammonis; 20, gyri of Andreas Retzius; 21, fimbria (displaced upward, arrows); 22, uncal apex; 23, band of Giacomini; 24, uncinate gyrus; 25, anterior perforated substance; 26, anterior commissure; 27, fornix; 28, corpus callosum. |
The Hippocampal Body
The hippocampal body, like the hippocampal head, is divided into intraventricular and extraventricular segments. It is bordered medially by the fimbria and laterally by the collateral eminence, which marks the intraventricular protrusion of cortex covering the collateral sulcus (Fig. 4.3). With the exception of the hippocampal head, the entire intraventricular hippocampal surface is covered by the choroid plexus. The extraventricular part of the hippocampus is limited to the dentate gyrus, fimbria, and superficial hippocampal sulcus (Fig. 4.1).
The Hippocampal Tail
The intraventricular part of the hippocampal tail is a transverse bulge. Digitations similar to those of the head are present on the tail’s internal surface, but they are not typically found on its intraventricular surface.
HISTOLOGY OF THE HIPPOCAMPUS
The hippocampus is a bilaminar archicortical (allocortex) structure consisting of Ammon horn (hippocampus proper) and the dentate gyrus, with one lamina rolled up inside the other (Fig. 4.3). The two internal structures are interlocked forming two U-shaped laminae that are separated from each
other by the hippocampal sulcus (Fig. 4.4), which is divided into a vestigial (deep) sulcus (10) and a superficial sulcus that is clearly visible on the mesial temporal lobe surface.
other by the hippocampal sulcus (Fig. 4.4), which is divided into a vestigial (deep) sulcus (10) and a superficial sulcus that is clearly visible on the mesial temporal lobe surface.
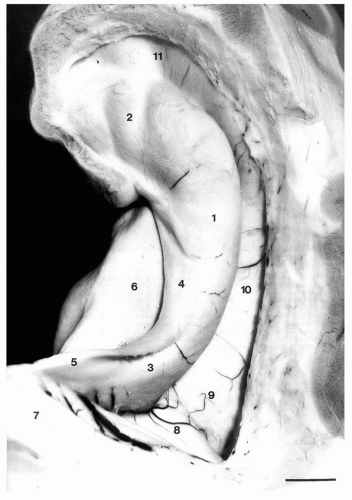 Figure 4.2 Intraventricular aspect of the hippocampus. The temporal horn has been opened and the choroid plexuses removed. 1, Hippocampal body; 2, head and digitationes hippocampi (internal digitations); 3, hippocampal tail; 4, fimbria; 5, crus of fornix; 6, subiculum; 7, splenium of the corpus callosum; 8, calcar avis; 9, collateral trigone; 10, collateral eminence; 11, uncal recess of the temporal horn. Scale bar: 6.5 mm. |
Hippocampal Layers
Ammon horn (cornu ammonis [CA]) may be divided into six layers: alveus, stratum oriens, stratum pyramidale, stratum radiatum, stratum lacunosum, and stratum moleculare (Fig. 4.5). These six layers are grouped into three allocortex layers: stratum oriens, stratum pyramidale, and stratum moleculare, which contains the last three layers (11,12).
The alveus covers the intraventricular surface and contains efferent hippocampal and subicular axons that enter the fimbria (Fig. 4.6), as well as afferent fibers mainly from the septum.
The stratum oriens, which has poorly defined borders, contains basket cells and is crossed by axons of pyramidal neurons to the alveus.
The stratum pyramidale contains the pyramidal cells, which are the major neurons of the hippocampus. From the base of these cells, axons connecting to the septal nuclei or association fibers traverse the stratum oriens toward the alveus. Pyramidal neurons have basal and apical dendrites (13). The basal dendrites arborize in the stratum oriens, whereas the apical dendrites traverse the cornu ammonis to reach the stratum moleculare in the proximity of the vestigial hippocampal sulcus. The pyramidal cells are surrounded by dense arborizations originating from basket cells in the stratum oriens (Fig. 4.7). The stratum pyramidale contains scattered basket cells (interneurons) and stellate cells (14).
The stratum radiatum contains mainly pyramidal cell apical dendrites that in CA1 are connected by the axons of the Schaffer collaterals, as well as septal and commissural fibers (Fig. 4.7).
The stratum lacunosum contains primarily perforant fibers from the entorhinal cortex and a lesser number of Schaffer collaterals (Fig. 4.7).
The stratum moleculare contains fewer interneurons and has broad arborizations of the apical dendrites from the pyramidal cells. It fuses with the stratum moleculare of the dentate gyrus as the vestigial hippocampal sulcus disappears during development.
Hippocampal Subfields
The CA contains four subfields, CA1 through CA4 (12), which are categorized according to the shape and location of the pyramidal neurons (Fig. 4.8).
CA1 neurons are triangular and multilayered and vary in sizes. CA1 (Sommer sector) (15) neurons continue into the five-layered subicular complex. CA2 neurons are larger, densely packed pyramidal cells. CA3 neurons are similar to CA2 neurons but are less densely packed. The CA3 subfield constitutes the genu of the CA. The CA3 subfield contains the mossy fibers, which are fine nonmyelinated fibers that originate from the granule neurons in the dentate gyrus and synapse with the stratum lucidum of CA3. CA4 neurons are large and triangular. Few in number, they are interspersed between the mossy fibers.
Dentate Gyrus (Fascia Dentata)
A concave structure on coronal sections that engulfs the CA4 subfield (Fig. 4.6), the dentate gyrus is separated from the hippocampal subfields by the vestigial hippocampal sulcus. It contains three layers that are more clearly identified that those of the CA: the stratum moleculare (long dendrites), stratum granulosum (granule cells), and the polymorphic layer or the subgranular zone, which contains mostly inhibitory interneurons of varied sizes (Figs. 4.7 and 4.8).
The polymorphic layer contains crossing axons that unite the neurons of the granular layer with those of CA4 and with interneurons in the polymorphic and CA4 regions.
The stratum granulosum is the main layer of the dentate gyrus and contains densely packed, small, round neurons. The stratum moleculare contains dendrites extending to the hippocampal fissure and a few interneurons. The outer and middle molecular zones receive fibers from the perforant pathway.
The stratum granulosum is the main layer of the dentate gyrus and contains densely packed, small, round neurons. The stratum moleculare contains dendrites extending to the hippocampal fissure and a few interneurons. The outer and middle molecular zones receive fibers from the perforant pathway.
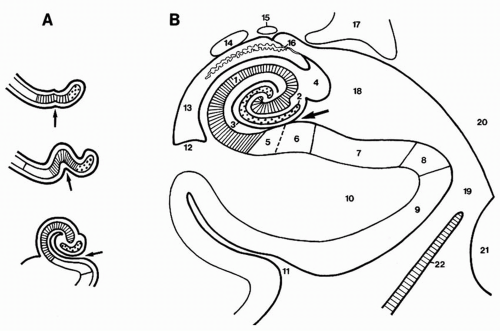 Figure 4.3 A: Development of the gyrus dentatus (dotted area) and of the cornu ammonis (hatched area) toward (B), their definitive disposition. Arrows indicate the hippocampal sulcus (superficial part). 1, Cornu ammonis; 2, gyrus dentatus; 3, hippocampal sulcus (deep or vestigial part); 4, fimbria; 5, prosubiculum; 6, subiculum proper; 7, presubiculum; 8, parasubiculum; 9, entorhinal area; 10, parahippocampal gyrus; 11, collateral sulcus; 12, collateral eminence; 13, temporal (inferior) horn of the lateral ventricle; 14, tail of caudate nucleus; 15, stria terminalis; 16, choroid fissure and choroid plexuses; 17, lateral geniculate body; 18, lateral part of the transverse fissure (wing of ambient cistern); 19, ambient cistern; 20, mesencephalon; 21, pons; 22, tentorium cerebelli. (Adapted from Duvernoy HM. The human hippocampus: functional anatomy, vascularization and serial sections with MRI. Berlin: Springer-Verlag, 1998, with permission.) |
Parahippocampal Structures
The subiculum is the continuation of the hippocampus from CA1. It is divided into the prosubiculum, subiculum proper, presubiculum, and parasubiculum.
HIPPOCAMPAL CONNECTIONS
The entorhinal area is the principal input to the hippocampus (Fig. 4.7). Composed of the periallocortex, it is divided into deep and superficial layers (16,17). The intrahippocampal circuits are divided into the polysynaptic and direct intrahippocampal pathways.
The Polysynaptic Pathway
The polysynaptic pathway (perforant pathway) originates in the layer II stellate neurons of the entorhinal cortex. Large pyramidal cells also contribute to the granular aspect of the cortical surface (Fig. 4.6) (18). Excitatory (glutamatergic) fibers reach the stratum moleculare of the dentate gyrus where they synapse with the dendrites of the granule cells (19). The axons of the granule cells (mossy fibers) project to the dendrites of the CA4 and CA3 neurons (20), but do not extend into CA2. These axons contain a high concentration of zinc (21,22). The axons of CA3 and CA4 cells project as the Schaffer collaterals to the apical dendrites of CA2 and CA1 cells (Fig. 4.7). A small number of axons project rostrally to the septal nuclei (Fig. 4.6). The main output of the hippocampus occurs caudally through CA1 axons that may directly project to the alveus and fimbria or indirectly project to the subiculum, and from there to the alveus and fimbria. The subiculum is the major hippocampal output relay area. All the neurons of the polysynaptic pathways are excitatory (glutamatergic) (23).
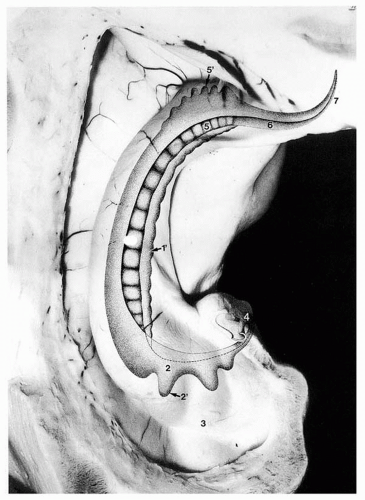 Figure 4.4 Gyrus dentatus seen through the hippocampus (transparent). 1, Gyrus dentatus in the hippocampal body; 1, margo denticulat us; 2, gyrus dentatus in the hippocampal head; 2′, digital extensions of the gyrus dentatus; 3, digitationes hippocampi; 4, band of Giacomini and terminal part of the gyrus dentatus in the medial surface of the uncus; 5, gyrus dentatus in the hippocampal tail; 5′, digital extensions of the gyrus dentatus; 6, fasciola cinerea; 7, terminal part of the fasciola cinerea. |
The rostral output of the polysynaptic pathway to the cortex is through the fornix to the mamillary bodies, reaching the anterior thalamic nucleus (directly or indirectly through the mamillary bodies) (24). The cingulate area and the retrosplenial cortex are reached from the thalamus (Fig. 4.9).
The input to the polysynaptic pathway is through fibers that originate in the parietal association area (area 7) and surrounding temporal and occipital areas (areas 40, 39, and 22). These fibers project to the entorhinal cortex through the parahippocampal gyrus. The polysynaptic pathway may be involved in episodic and spatial memory, the most primitive memory types (25).
Direct Intrahippocampal Pathway
The direct intrahippocampal pathway originates in layer III of the entorhinal gyrus. These fibers reach directly to the pyramidal neurons of CA1, from which axons project directly to the subiculum and then to the deep layers of the entorhinal cortex (Fig. 4.10) (26,27). The input to the direct intrahippocampal pathway is from the inferior temporal association cortex. The output of the pathway is to the temporal association cortex, the temporal pole, and the prefrontal cortex (Fig. 4.11). The direct pathway may be involved in semantic memory (25).
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


