Chapter 23 Intervertebral Disc
Anatomy, Physiology, and Aging
Anatomy
Intervertebral Disc
The spinal column has 23 IVDs, starting from the C2-3 interspace to the L5-S1 interspace. These discs constitute approximately 25% of the total height of the spinal column. The discs vary in thickness from the thinnest disc in the thoracic region to the thickest in the lumbar region.1 The IVD is a vital component of the joint system present at each spinal level. This system allows for the range of motion permitted at each segment.2 Each IVD works in conjunction with the paired dorsal zygapophyseal joints to form a “three-joint complex.” The function of the individual components of this complex are intimately related, as are their effects on one another during the degenerative process.
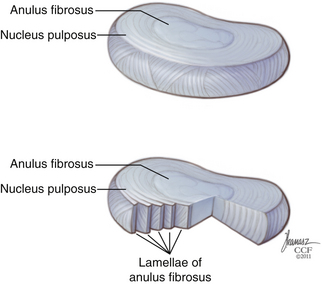
The IVD is constructed of three distinct components. The bulk of the disc is made up of a central nucleus pulposus (NP) and the surrounding anulus fibrosus (AF) (Fig. 23-1). The disc is flanked rostrally and caudally by cartilaginous end plates that serve as a transitional zone between the disc and the adjacent vertebral bodies.3
Nucleus Pulposus
The central core of the IVD is composed of the NP. It is formed from remnants of the notochord, a derivation of the endoderm—unlike the remaining components of the IVD, which are derived from the mesoderm.4 The NP is composed of a soft gelatinous material consisting of proteoglycans, surrounded circumferentially by the AF. The NP is 80% water, with proteoglycans contributing to 50% of its dry weight. Aggrecans, a type of proteoglycan known as leucine-rich repeat proteins, are the predominant type of proteoglycans in the NP.5 The high-proteoglycan content allows for the NP’s ability to maintain an increased hydration state, which creates the viscoelastic properties of the NP, which are essential to its load-bearing properties in the spine. The NP is rich in type II collagens, which constitute 80% of the collagen content in the NP.6 There is a clear distinction between the NP and the surrounding AF; however, this distinction becomes less obvious during the course of aging.1 Elastin fibers can also be found in the NP. These fibers are observed to be situated in both a radial distribution from the center to the periphery in the NP, as well as in a vertical orientation, anchoring the NP to the end plates.7 This orientation likely contributes to maintaining the structure of the NP within the AF, restoring the NP to its original form following load bearing, as well as playing a role in load transmission to surrounding AF.
Anulus Fibrosus
The AF is a concentrically organized structure that surrounds the NP, occupying the majority of the disc space. It is composed of bundles of collagen fibers that are arranged in concentric lamellae. Approximately 15 to 25 lamellae compose each anulus.8 The collagen fiber bundles within the lamellae are generally oriented at 30 degrees from the horizontal axis, although closer examination shows that throughout the course of the lamellae, the fiber angle can vary from 20 to 55 degrees.9 Collagen fibrils in adjacent lamellae run in the opposite direction, allowing for greater tensile strength during stretching.
The AF can be further separated into an outer and inner anulus. The outer AF is a tougher, less flexible component, composed primarily of densely packed type I collagen fibers. The cells in the outer AF are more ellipsoidally shaped cells and are fibroblast-like in nature. At the periphery of the outer anulus, fibrillar bundles known as Sharpey fibers extend superiorly and inferiorly to anchor the disc into the periosteal fibrils of the adjacent vertebral bodies.10 The inner AF is a softer, less dense, and more fibrocartilagenous component, composed primarily of type II collagen fibers. It possesses a less structured cellular morphology with more widely spaced cells than that of type I collagen. Type II collagen has been shown to be able to maintain 50% to 100% greater water content than type I collagen.11 The outer lamellae are composed of up to approximately 70% type I collagen and only 20% type II collagen, whereas in the inner AF the percentage of type II collagen increases to 70%. There is also a progressive increase in the concentration of proteoglycans from 10% to 30% when moving from the outer to inner AF.12 Other types of collagen are also present in the anulus, with type VI collagen making up as much as 10% of the collagen in the AF.5
Cartilage End Plates
On the superior and inferior surfaces of the IVD lies the cartilage end plates. They are composed of a thin layer of hyaline cartilage, which serves as the interface between the bony vertebral body and the disc itself. Their composition is similar to the disc; that is, made up of proteoglycans, type II collagen, and water.13 The end plates range from 0.5 to 1.5 mm in thickness. They are thinnest at the center where they interface with the NP.14 Each end plate is attached to the adjacent vertebral body by a thin layer of calcium. The calcium is absent in areas where numerous perforations exist throughout the end plates. These perforations in the end plates allow for the passage of vascular channels that traverse from the adjacent vertebral body into the disc.1 These channels become progressively rare and eventually nonexistent, as the disc ages past the second decade of life.15.The end plate serve as a stiff but porous barrier between the vertebral body and the IVD. They allow for the diffusion of nutrients and fluid movement into and out of the disc and prevent protrusion of the soft malleable disc material into the vertebral body (Schmorl nodes). Weaknesses in the end plates resulting in Schmorl nodes may be due to a dysregulation of the end-plate composition since those areas have been found to have significantly less proteoglycan concentration.16
Vascular Supply
The IVD is generally considered to be an avascular tissue structure, but this is only partially accurate. In infancy, the native disc has a direct vascular supply. This, however, nearly completely regresses by the third decade of life. Vascular “buds” extend from the vertebral bodies through the porous channels in the cartilage end plates and traverse into the disc to supply nutrients.16 These channels are overtaken by scarring or collapse, likely as a result of weight bearing, past the second decade of life and become occluded and eventually nonexistent.15,17 Without a direct vascular supply, nutrient exchange occurs through diffusion from blood vessels that surround the periphery of the anulus. The surrounding blood supply comes from segmental branches of the spinal artery.18
Innervation
The IVD receives innervations primarily from two sources: the sinuvertebral nerve and the sympathetic trunk via the multiple gray rami communicantes (Fig. 23-2). The sinuvertebral nerve is formed from a branch of the ventral primary ramus and the gray ramus communicans, which branches from the sympathetic trunk. The sinuvertebral nerve enters the spinal canal caudal to the pedicle through the intervertebral foramen and branches into a larger superior division and a lesser inferior division. The superior division travels lateral to the posterior longitudinal ligament in the ventral aspect of the spinal canal, supplying sensation to the dorsal and dorsolateral anulus, as well as the posterior longitudinal ligament. The inferior trunk passes medially and caudally. The lateral and ventral portions of the AF are supplied by multiple gray rami communicantes from the nearby sympathetic trunk.19,20 The sympathetic trunk runs in parallel, ventral and lateral to the vertebral column, composed of nerve root contributions from the thoracic and upper two segments of the lumbar spinal cord. The sympathetic trunk provides a number of gray rami comminicantes that help supply the anulus. At least one, but often more, rami communicantes travel around the vertebral body deep to the psoas muscle to join with each ventral primary ramus. Rami communicantes also traverse the psoas to join with the ventral primary ramus. In addition, “paradiscal” rami run along the surface of the IVD in the perianular connective tissue.19 Innervation of the end plate extends from intraosseous nerves that branch from the ventral primary rami and the basivertebral nerve, a branch of the sinuvertebral nerve, which enters the end plate dorsally.6
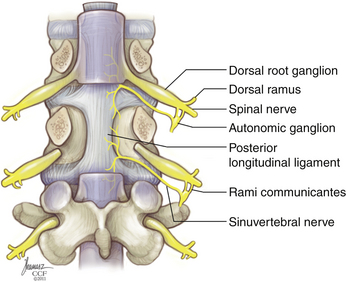
Innervation of the disc interspace is restricted to the outer anulus. A quantitative analysis of nerve density in multiple IVDs showed innervation to extend to a depth of four to seven lamellae ventrally and no more than three lamellae dorsally. This is indeed very shallow, given that a usual AF contains an average of 15 to 25 lamellae.8 Unlike the anulus, innervation of the end plate is densest near the center, adjacent to the NP. Innervation of the end plates originates from the intraosseous nerves that penetrate the caudal portion of the vertebral body ventrally and dorsally and travel to the center of the end plate.21
Physiology
Extracellular Matrix
The main purpose of the IVD is to serve as a load-bearing structure. The disc performs this task by absorbing axial loads and redistributing them across the entire disc. The capacity for load redistribution is determined, in part, by the molecular structure of the disc. The extracellular matrix is primarily composed of collagen and proteoglycan. These molecules allow the different components of the disc to complete the aforementioned tasks. As previously noted, the AF is a mix of type I and type II collagen, which gives it its necessary tensile strength. In addition to type I and II collagen, multiple other collagen types, including types V and XI, are present in smaller amounts. These collagens play a role in interlinking the collagen fibrils, thus contributing to the overall strong fibrillar collagen network.5,22 The differences in the composition in different areas of the disc create a dynamic structure that is able to bear and distribute loads across the entire structure. When axial loading takes place, the NP and inner AF absorb the weight and conform to generate hydrostatic pressure that is distributed evenly to the adjacent outer anulus. The alternating lamellar structure of the type I collagen network in the outer AF creates the tensile strength to absorb the redistributed loading force.
Proteoglycans are the other main molecular component of the disc. They constitute 50% of the cells in the NP. Numerous types of proteoglycans are present in the extracellular matrix, including aggregan, versican, decorin, biglycan, fibromodulin, lumican, and perlecan. Aggregan is the most important proteoglycan found in the NP. Proteoglycans comprise a central core protein with attached side chains of keratin sulfate and chondroiton sulfate. Early in life, aggrecans are rich in chondroitin sulfate chains, but these side chains are gradually replaced by keratin sulfate as the disc matures. At their N-terminus, aggrecans attach to hyaluronic acid,12 and at their C-terminus, they can attach to various molecules in the extracellular matrix, including collagen.23 The proteogylycans are attached to hyaluronic acid molecules through link proteins, which allow them to form aggregates.5 Proteoglycans have negative charges on their surface and are therefore hydrophilic molecules able to bind water—thus allowing the NP to retain its hydrated state. The percentages of proteoglycan aggregates are highest in infancy and decrease with age. Breakdown of aggregans that occurs with aging causes them to be replaced by nonaggregated proteoglycans, which are less able to absorb water than their aggregated counterparts.
Related posts:
 Definition and Assessment of Dysfunctional Segmental Motion
Pathophysiology of Cervical Myelopathy: Biomechanics and Deformative Stress
Cervical Skip Corpectomy
Percutaneous and Minimally Invasive Approaches to Decompression and Arthrodesis of the Thoracolumbar Spine
Medical Management of the Patient with Acute Spinal Cord Injury
Spine Infection
Definition and Assessment of Dysfunctional Segmental Motion
Pathophysiology of Cervical Myelopathy: Biomechanics and Deformative Stress
Cervical Skip Corpectomy
Percutaneous and Minimally Invasive Approaches to Decompression and Arthrodesis of the Thoracolumbar Spine
Medical Management of the Patient with Acute Spinal Cord Injury
Spine Infection
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


