 We first came across the concept of the hierarchical structure of the brain, its organization in an ascending series of levels, in Chapter 9. Now, in this final chapter we look at the very highest levels of all, those that determine what we do rather than how we do it.
We first came across the concept of the hierarchical structure of the brain, its organization in an ascending series of levels, in Chapter 9. Now, in this final chapter we look at the very highest levels of all, those that determine what we do rather than how we do it.local gradients, the path it traces out being a direct function of its environment. Very simple tropistic mechanisms like these give rise to surprisingly life-like behaviour, as in Dr Grey Walter’s pioneering electromechanical tortoise Elsie which ran amiably around the floor looking for light, or the interactive version in NeuroLab.
 Though higher animals produce more complex behaviour, the mechanism is essentially the same. The added complexity comes about for two reasons.
Though higher animals produce more complex behaviour, the mechanism is essentially the same. The added complexity comes about for two reasons.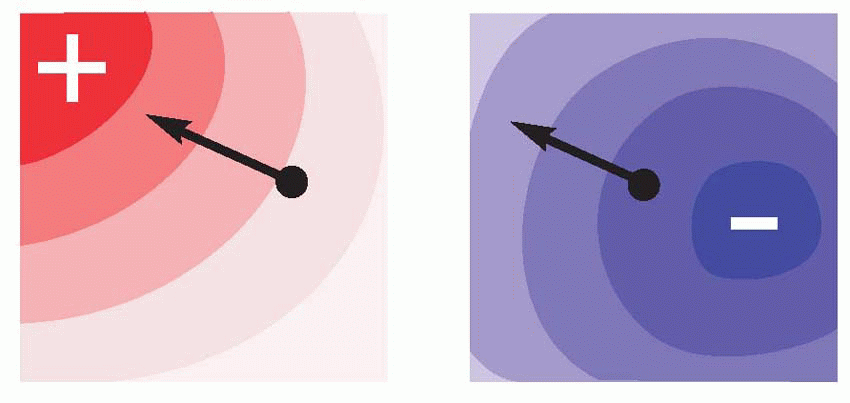 |
 |
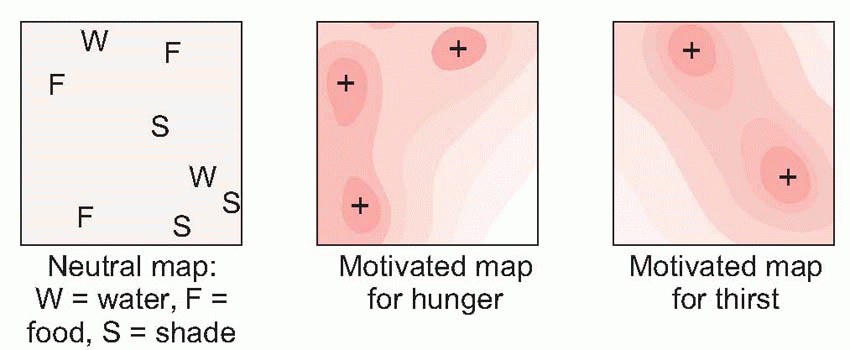 |
associated with a particular location. Similarly, brain scans of London-taxi drivers, who have to undergo a period of rigorous spatial learning, demonstrate an enlarged posterior hippocampus relative to matched controls.
 Equally, it is the hypothalamus that provides information about need, about the state of the body, and projects to the hippocampus via the septal nuclei. It is the centre to which autonomic afferents project, and its neurons monitor such physiological states of the blood as glucose concentration, temperature and osmolarity, as well as levels of circulating hormones, that decides one’s state of need. It is also in the hypothalamus that primary consummatory responses such as eating and drinking may be triggered off by electrical stimulation. The hypothalamus is thus utterly at the heart of the neural mechanisms that generate motivation: it is discussed in detail later in this chapter (p. 278).
Equally, it is the hypothalamus that provides information about need, about the state of the body, and projects to the hippocampus via the septal nuclei. It is the centre to which autonomic afferents project, and its neurons monitor such physiological states of the blood as glucose concentration, temperature and osmolarity, as well as levels of circulating hormones, that decides one’s state of need. It is also in the hypothalamus that primary consummatory responses such as eating and drinking may be triggered off by electrical stimulation. The hypothalamus is thus utterly at the heart of the neural mechanisms that generate motivation: it is discussed in detail later in this chapter (p. 278).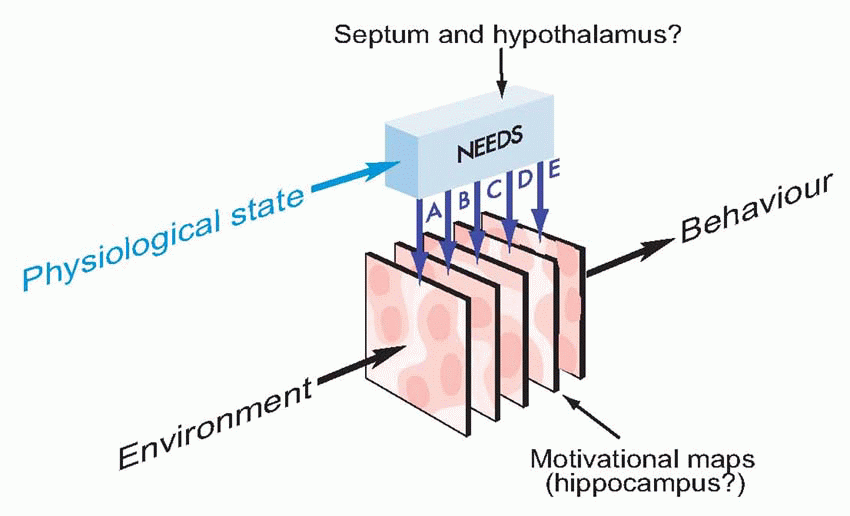 |
 |
 To some extent it is in fact possible to plot motivational maps in Man: by averaging over large numbers of individuals, it is not difficult to measure quite directly the same kinds of tropistic gradients for us humans, that work so well in describing what an amoeba does. If you take a group of people and ask them the very simple question ‘Where in Britain would you like to be?’, it is possible to obtain contour maps of average preferences, in this case of the relative desirability amongst a group of school-leavers of different parts of the country. These are certainly motivational maps, in the sense that if the individuals had the means to do it, they would be translated into actual migratory behaviour not very different in essence from our amoeba moving blindly down its tropistic gradient.
To some extent it is in fact possible to plot motivational maps in Man: by averaging over large numbers of individuals, it is not difficult to measure quite directly the same kinds of tropistic gradients for us humans, that work so well in describing what an amoeba does. If you take a group of people and ask them the very simple question ‘Where in Britain would you like to be?’, it is possible to obtain contour maps of average preferences, in this case of the relative desirability amongst a group of school-leavers of different parts of the country. These are certainly motivational maps, in the sense that if the individuals had the means to do it, they would be translated into actual migratory behaviour not very different in essence from our amoeba moving blindly down its tropistic gradient.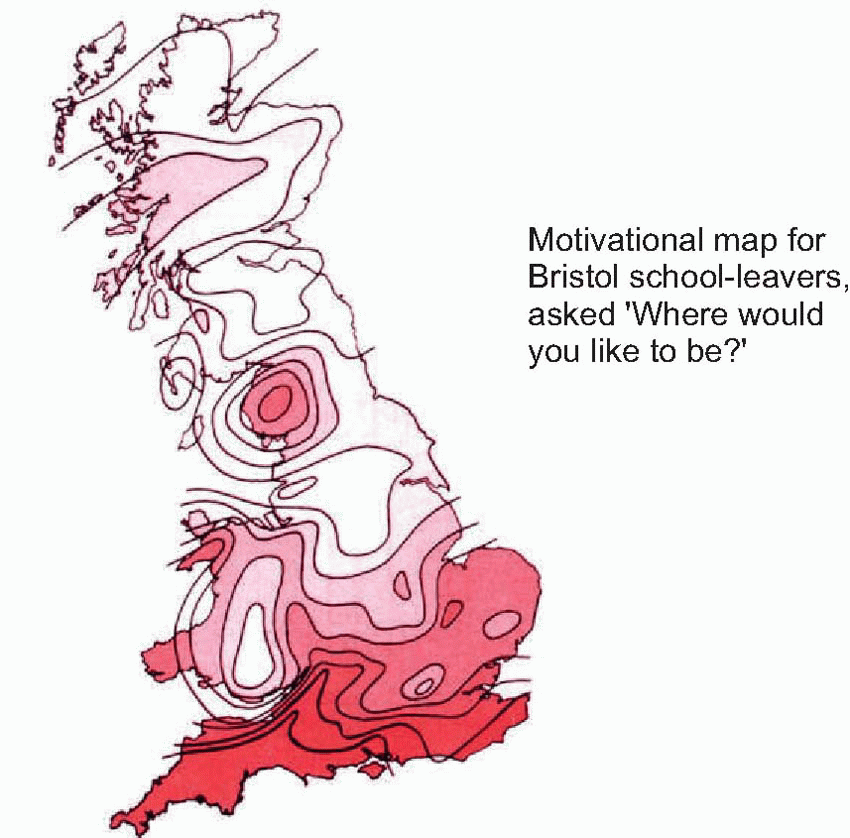 |
 |
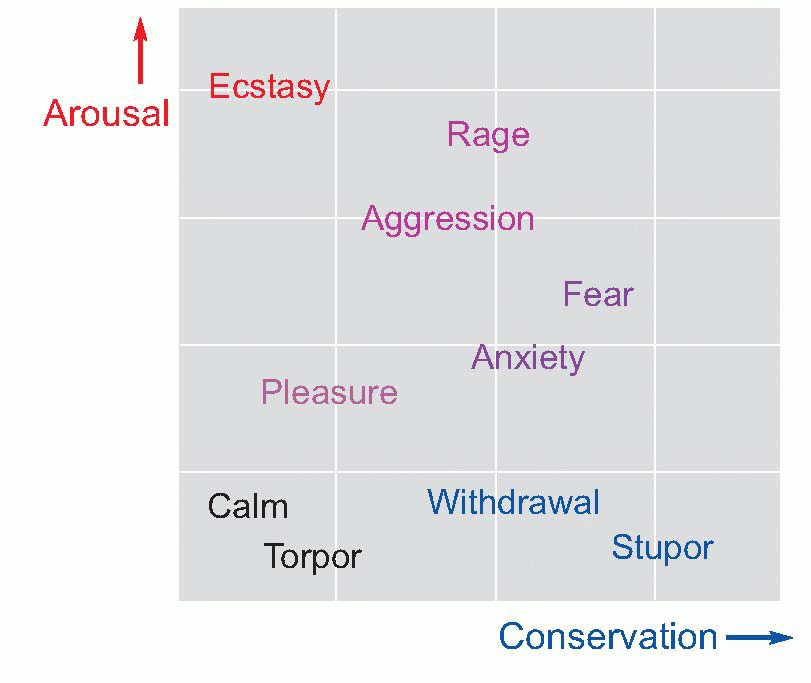
desirable goal or away from a source of threat (p. 274): the state often described by physiologists as ‘fight, fright or flight’, that results in an increase in the general activity of the sympathetic system, and the release of adrenaline. The consequent bodily responses are all of more or less obvious use in preparing the body for the expenditure of the energy that will be used to achieve the goal: blood flow through the muscles is increased, the heart rate is raised, glucose is released into the blood, the bronchioles and pupils dilate, the electrical activity of the brain increases, reaction times get quicker, and there is an associated feeling of general excitement. All of this of course involves a certain expenditure of energy, and would be a drain on the body’s resources if kept up for a long time: but much is now at stake, and the gamble is one worth taking.
 |
 |
rarely a frightening experience in modern society – but rage can easily be elicited in situations of frustration, when the positive and negative aspects of a possible goal are nicely balanced. The nature of the goal clearly affects the precise response that is made, yet there can often be a curious generality about arousal, most obvious perhaps in human sexual behaviour: sexual aggression shades off imperceptibly into sadism, and affection is expressed by licking and biting and other responses more appropriate to an edible goal.
 |

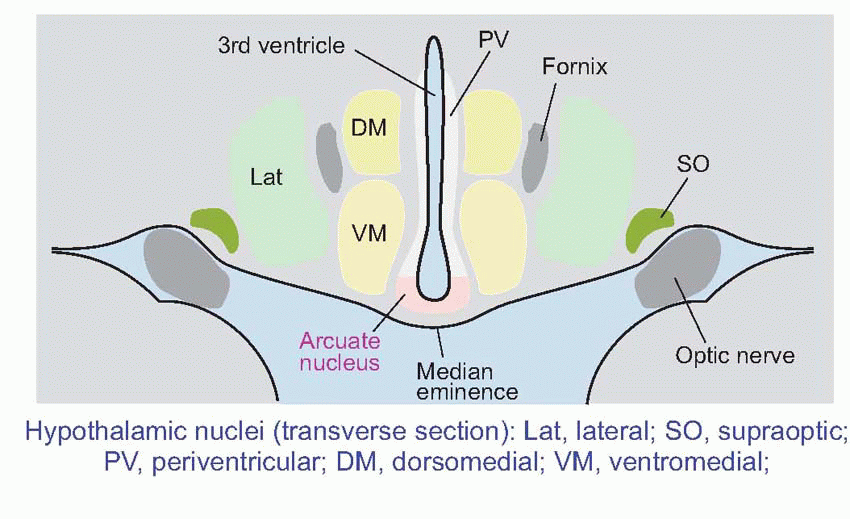
 The hypothalamus lies on either side of the third ventricle, immediately above the pituitary (hypophysis) and below the thalamus, and consists of several fairly distinct nuclei. At its lowest level, it controls the pituitary (hypophysis), through two distinct mechanisms: one is direct, the other indirect.
The hypothalamus lies on either side of the third ventricle, immediately above the pituitary (hypophysis) and below the thalamus, and consists of several fairly distinct nuclei. At its lowest level, it controls the pituitary (hypophysis), through two distinct mechanisms: one is direct, the other indirect.
Direct: the axons of magnocellular neurons (black, p. 279) in the supraoptic and paraventricular nuclei pass right down into the pituitary stalk to terminate in the posterior lobe (neurohypophysis). Here they release their transmitters, not at synaptic junctions but directly into the bloodstream; thus these neurons are acting directly as endocrine cells, and their transmitters are actually hormones. Neurons of the supraoptic region predominantly release antidiuretic hormone (ADH), those of the paraventricular region mostly oxytocin. Both these hormones are nonapeptides of very similar structure, but their effects are entirely different. ADH helps control the osmolarity of the blood by stimulating the retention of water in the kidney; in large doses it may also increase blood pressure through arteriolar constriction (hence its alternative name of vasopressin). Oxytocin stimulates the smooth muscle of the uterus in labour, and also causes milk ejection during lactation; in both cases the stimulus to its release is essentially neural, predominantly from mechanoreceptors in the regions concerned.
 |
Indirect: the other route by which the hypothalamus controls the secretion of hormones from the pituitary is quite different. Axons from other, parvocellular, hypothalamic neurons terminate in a region on the ventral surface (the median eminence) where a system of fenestrated capillaries carries arterial blood down to the anterior pituitary through a portal system. The substances released from their terminals (releasing or inhibiting hormones: black dots) enter this portal system and are transported to the anterior pituitary where they each either stimulate or inhibit the release of some corresponding pituitary hormone from endocrine cells (blue). Thus the release of the pituitary hormone prolactin, which stimulates the secretion of milk and has other functions related to pregnancy, is stimulated by prolactin-releasing hormone (PRH) and inhibited by prolactin-inhibiting hormone (PIH), from medial regions of the hypothalamus. Other hypothalamic hormones have their corresponding pituitary ones: apart from PIH (which is known to be dopamine) they are all small peptides.
The substances released from their terminals (releasing or inhibiting hormones: black dots) enter this portal system and are transported to the anterior pituitary where they each either stimulate or inhibit the release of some corresponding pituitary hormone from endocrine cells (blue). Thus the release of the pituitary hormone prolactin, which stimulates the secretion of milk and has other functions related to pregnancy, is stimulated by prolactin-releasing hormone (PRH) and inhibited by prolactin-inhibiting hormone (PIH), from medial regions of the hypothalamus. Other hypothalamic hormones have their corresponding pituitary ones: apart from PIH (which is known to be dopamine) they are all small peptides.
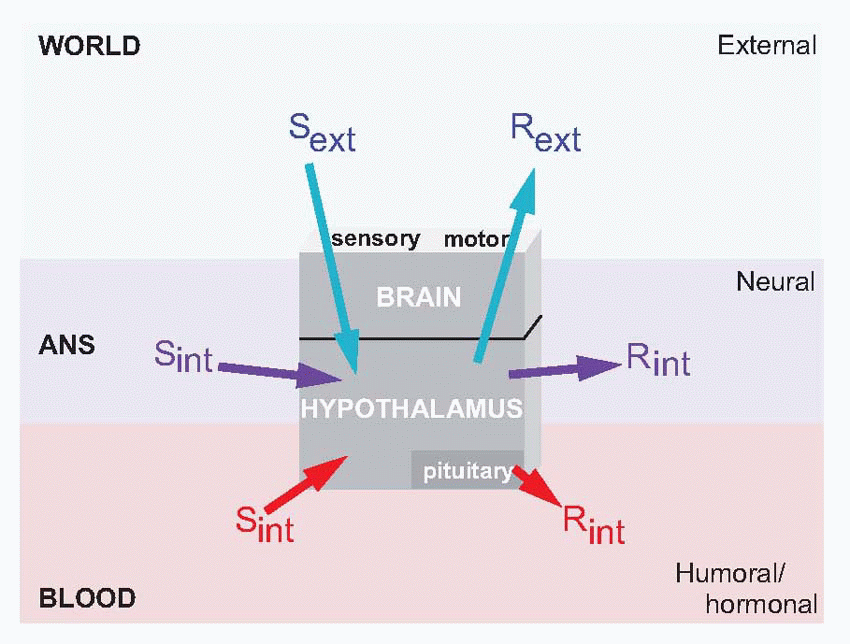
doing this it relies on the fact that it is itself capable of monitoring critical aspects of the blood, notably blood glucose, temperature and osmolarity: the hypothalamus is a death predictor. At an intermediate level it acts as what Sherrington called the head ganglion of the autonomic system, effectively being in charge of the whole of the milieu intérieur, through its autonomic efferents that control the heart, digestive tract and other vital organs. Again, it is able to do this effectively because it has feedback from the body through autonomic afferents. In both cases, sensory information about the internal state of the body – Sint – is used as feedback to create homeostatic internal responses – Rint. The intermingled coordination of these two kinds of feedback control – partly hormonal, partly neural – is what keeps us alive.
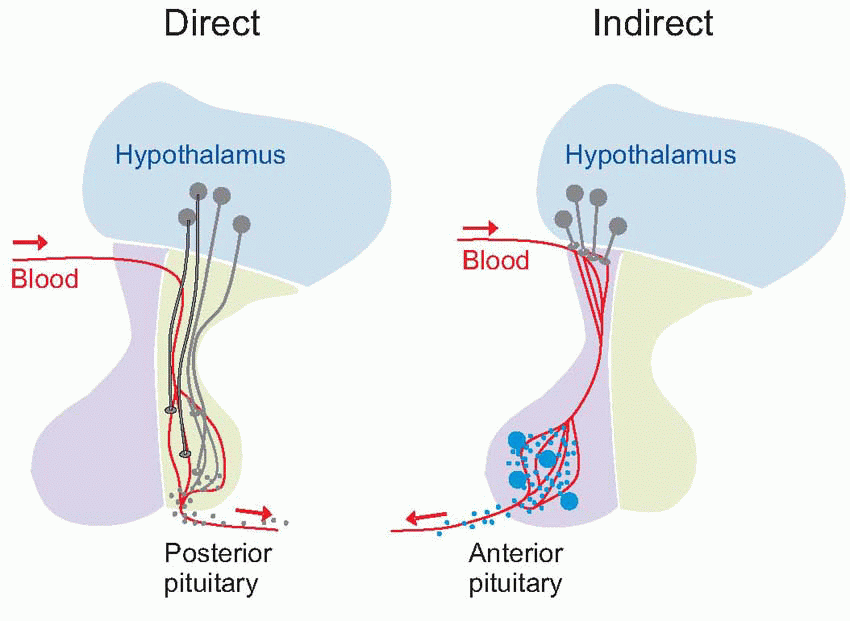 |
Table 14.1 Hypothalamic control of the pituitary | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
response to a river if you’re thirsty is to bend down and drink. The hypothalamus is both the ultimate detector of need, and the ultimate generator of behaviour: the rest of the brain is really just a way of making the hypothalamus work better.
 |
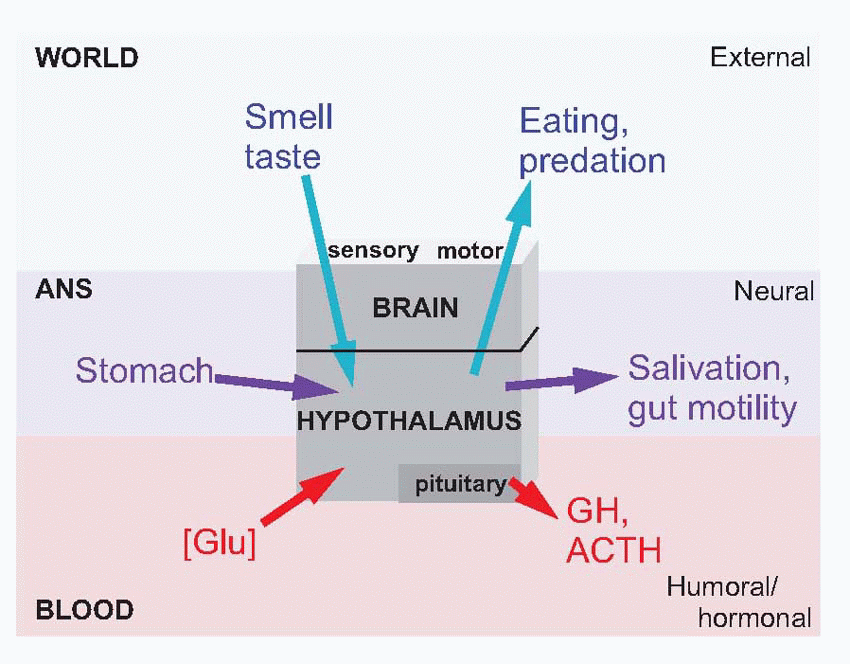 |

through overt behaviour such as curling up in the cold and seeking warmth (not to mention putting on or taking off one’s clothes). Once again, the input to this system is partly neural and partly humoral; afferent signals from somatosensory warm and cold receptors, and from cells in the anterior hypothalamus that themselves respond to the temperature of the blood. Temperature regulation appears to be represented rather diffusely in the hypothalamus. Electrical stimulation at many points can produce fragments of temperature-regulating activity such as shivering, piloerection, vasoconstriction and sweating. Broadly speaking, the anterior half is concerned with mechanisms for losing heat in a hot environment and the posterior half with conserving heat when it is cold. More generally, there is a tendency for sympathetic responses to be found in the posterior half and parasympathetic in the anterior half. The fact that none of these effects is sharply localized simply reflects the high degree of inter-relationship that exists between different homeostatic functions: a given response such as vasoconstriction may be caused by many diverse kinds of stimulus (fear, low temperature, low blood pressure), and will in turn have a disturbing effect on several different homeostatic systems. The thyroid-stimulating hormone is controlled by TRH, associated with more ventral parts of the hypothalamus. The thyroid hormones have many interrelated effects on metabolism and growth, which are not well understood. One of its functions appears to be to cause a general increase in metabolic rate, helping to maintain body temperature under conditions of chronic cold.
 The ancient neural pathways to and from the hypothalamus are not well understood. In particular, although it is evident that the hypothalamus is indeed ‘the head ganglion of the autonomic system’, it has not been possible to identify conclusively by what routes its autonomic functions are mediated. Its connections with the limbic system are clearer: it receives afferents from the hippocampal region through a massive fibre bundle (the fornix), and is interconnected through the medial forebrain bundle with many parts of the limbic system including the amygdala, with orbitofrontal cortex, and with reticular formation (RF), including the areas controlling respiration and the cardiovascular system (see p. 204). The medial forebrain bundle also carries afferent olfactory information. Some hypothalamic nuclei – notably the supraoptic and paraventricular, project to extraordinarily diverse areas, including substantia nigra and the substantia gelatinosa of the dorsal horn. Corresponding to this diverse output, stimulation of many regions of the hypothalamus can give rise to fragments of emotional behaviour such as sweating, piloerection, freezing, sexual responses, and so on.
The ancient neural pathways to and from the hypothalamus are not well understood. In particular, although it is evident that the hypothalamus is indeed ‘the head ganglion of the autonomic system’, it has not been possible to identify conclusively by what routes its autonomic functions are mediated. Its connections with the limbic system are clearer: it receives afferents from the hippocampal region through a massive fibre bundle (the fornix), and is interconnected through the medial forebrain bundle with many parts of the limbic system including the amygdala, with orbitofrontal cortex, and with reticular formation (RF), including the areas controlling respiration and the cardiovascular system (see p. 204). The medial forebrain bundle also carries afferent olfactory information. Some hypothalamic nuclei – notably the supraoptic and paraventricular, project to extraordinarily diverse areas, including substantia nigra and the substantia gelatinosa of the dorsal horn. Corresponding to this diverse output, stimulation of many regions of the hypothalamus can give rise to fragments of emotional behaviour such as sweating, piloerection, freezing, sexual responses, and so on. The amygdala (‘almond’ – from its shape) appears to be an important structure on the output side: it projects partly to the hypothalamus (generating autonomic and hormonal effects) and is also capable of generating actual behaviour by two routes to the motor system: to dorsomedial nucleus of the thalamus, which projects in turn to frontal cortex; and to the nucleus accumbens (ventral striatum) which projects to parts of basal ganglia, particularly globus pallidus and substantia nigra. It can be broken down into three major divisions (basolateral, cortical and centromedial), further subdivided into 10 or so nuclei. The projection from the centromedial region to
The amygdala (‘almond’ – from its shape) appears to be an important structure on the output side: it projects partly to the hypothalamus (generating autonomic and hormonal effects) and is also capable of generating actual behaviour by two routes to the motor system: to dorsomedial nucleus of the thalamus, which projects in turn to frontal cortex; and to the nucleus accumbens (ventral striatum) which projects to parts of basal ganglia, particularly globus pallidus and substantia nigra. It can be broken down into three major divisions (basolateral, cortical and centromedial), further subdivided into 10 or so nuclei. The projection from the centromedial region to the hypothalamus through the medial forebrain bundle is particularly interesting, as its organization seems to correspond rather nicely with the two fundamental types of emotion described earlier in this chapter, conservation and arousal.
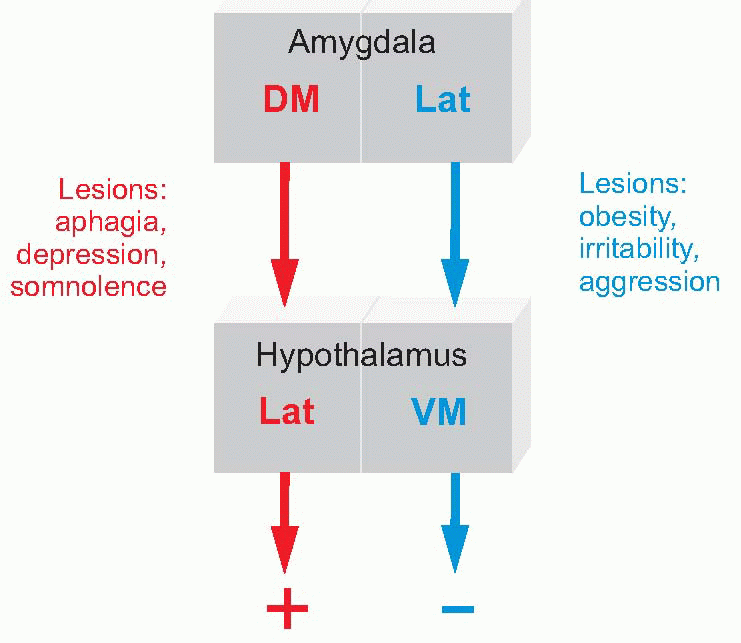 |
The lateral area projects directly to the ventromedial hypothalamus, and broadly speaking both seem to correspond with conservation, the horizontal axis of the diagram on p. 277. Stimulation in this region of the amygdala produces passivity, and even sleep or stupor. Lesions in the ventromedial hypothalamus, the ‘satiety centre’, produce over-eating and obesity; again, similar effects are produced in the lateral amygdala, but with a more general affective change as well, an increase in irritability and aggressiveness.
The dorsomedial area projects directly on to the lateral hypothalamus, and both regions seem, broadly speaking, to be concerned with arousal, the activation of a positive motivational drive. We have already seen that the lateral hypothalamus is associated with the initiation of eating and drinking behaviour, in the sense that lesions give rise to aphagia and adipsia; but they also produce a more general depression: dogs with lateral hypothalamic lesions are described as having a sad appearance, and are listless and somnolent. Lesions of the dorsomedial amygdala have similar effects, with perhaps more of the general, affective, component; stimulation in this region may produce hissing and growling and other signs of positive arousal.
Large bilateral lesions that include the amygdala create an animal in which tropistic behaviour is greatly exaggerated (the Klüver-Bucy syndrome): everything in the environment seems indiscriminately attractive, and such a monkey will compulsively examine and try to eat such things as the bars of its cage and its own faeces, and even things like snakes that would terrify a normal animal. The same kind of hypertropism is seen in its sexual activity: the animal is markedly hypersexual and may try to copulate with members of its own sex, as well as inanimate objects. Again, it is as if objects do not receive their normal emotional colouring, and inappropriately directed behaviour results, though a complicating factor is that such widespread lesions also damage regions of temporal cortex involved more generally in visual recognition. Studies in humans have confirmed the general sense that the amygdala is in part involved in fear, aversion from stimuli that have been experienced as harmful. Early studies showed that electrical stimulation of the amygdala in conscious patients (as part of preliminary investigations of possible sites of epileptic disturbances) elicited feelings of fear, and lesions in this area are reported to interfere with the formation of associations with unpleasant stimuli. Some other studies suggest that the amygdala is concerned not just with aversive responses but also with positive responses to stimuli associated with reward.
 At the beginning of this chapter it was pointed out that the whole purpose of the evolutionary development of the brain is to help the hypothalamus in carrying out homeostasis, by making predictions: predictions about what we are about to experience, predictions about what
At the beginning of this chapter it was pointed out that the whole purpose of the evolutionary development of the brain is to help the hypothalamus in carrying out homeostasis, by making predictions: predictions about what we are about to experience, predictions about what will be the result of doing this rather than that. If the world behaved with clockwork regularity, and if we had unlimited information about it, making decisions about how to respond to a particular stimulus would be entirely trivial, like playing noughts-and-crosses. But that is not the case; and just as a successful business makes the best estimates of the risks and potential gains from taking one course of action rather than another, so that its gambles tend to pay off, so the neural mechanisms that determine what we do must similarly compute the probabilities of different outcomes, and the rewards likely to accrue from different actions. The rational way to proceed is to calculate, over all the things we might do, the one for which the expected return (the size of the reward multiplied by the probability of getting it) is maximum. This would be relatively straightforward, if our sensory systems were fast, reliable and capacious: but they are not. As we have seen again and again, they are subject to noise (some originating in the outside world, some internally), and – by computer standards – operate with a very low bandwidth. The result, as has been repeatedly emphasized, is that we are continually making guesses about what’s going on, based mostly on our experience of what has happened in the past (embodied in our internal models) and only partly on real information trickling in through our senses (p. 255). The situation is not entirely unlike making a clinical diagnosis: on the one hand, the doctor has a lifetime’s experience of the probability of different kinds of disease, and this is supplemented by the often rather meagre information afforded by the patient in front of them. The question then arises, how to combine these two kinds of information – actual evidence and prior expectation – in order to arrive at the most likely assessment of the underlying cause.
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


