5 Neurology of sensory and receptor systems
 Clinical cases for thought
Clinical cases for thoughtIntroduction
Sensory information is essential for a variety of reasons such as to construct a perception of ourselves in the universe, muscle movement control, internal organ and blood flow functionality, maintaining arousal, and developing and maintaining survival and plasticity in neural networks.
Reception and sensation of afferent stimulation
The physical properties of the stimuli detected by our receptors often differ dramatically from the perceptions that we form from these stimuli. For example, our retinal cells detect electromagnetic radiation patterns but we perceive the Mona Lisa. Our ears detect sound wave variations and patterns but we perceive a Mozart symphony. Our nociceptors detect chemical imbalances but we perceive pain. This amazing ability of our brain to process this information in an individually unique but universally characteristic fashion is one of the fundamental attributes that determines our humanism. Colours, sounds, feelings, pain, and all other human perceptions do not exist outside of the human mind. Thus our perception of our universe is not a true representation of our physical universe but a mental construct composed by our mind’s interpretation of all of the integrated nervous input available to it at any given moment (Martin & Jessell 1995). It is important to remember this when a patient presents with symptoms that cannot be explained by our traditional understanding of neuroscience or neuroanatomy. The patient’s perception is their reality. For example, in people with fibromyalgia, the detection of movement by their joint receptors is perceived as pain in their mind. To this patient this perception of pain is very real.
The sensory neurons
 Quick facts 5.1
Quick facts 5.1Somatic sensation
Somatic sensibility has four major types of modalities:
1. Discriminative touch—Discrimination of size, shape, texture, and movement across the skin.
2. Proprioception—The sense of static and kinetic position of the limbs and body without visual input.
3. Nociception—The signalling of tissue damage or chemical irritation, typically perceived as pain or an itch.
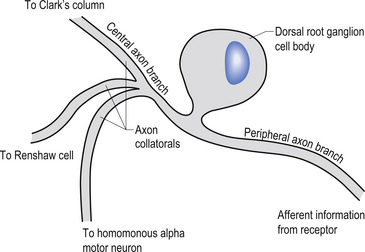
The cell bodies of the primary afferent neurons of the spinal cord live in the dorsal root ganglia situated adjacent to the spinal cord in the immediate vicinity of the intervertebral foramen from which they enter the spinal canal (Schwartz 1995). The primary afferent neurons are classified as pseudounipolar cells because they give rise to only one axon that transmits information from the periphery to the spinal cord. The segment of the axon that transmits information from the peripheral receptor towards the cell body is called the peripheral axon branch, and the portion of the axon transmitting information away from the cell body towards the spinal cord is called the central axon branch. The dorsal root neurons do not form dendrites as most other neurons do; thus any modulatory activity occurring at these neurons must occur at the cell body or presynaptically on the central process as it enters the grey matter spinal cord. Most of the central processes of the primary afferent develop collateral axon branches as they enter the spinal cord (Fig. 5.1). These collateral branches synapse with a variety of other neurons depending on the type of modality that the primary afferent cell is relying. The collateral branching and synapse formation of each type of primary afferent will be described in detail as each individual modality is described below.
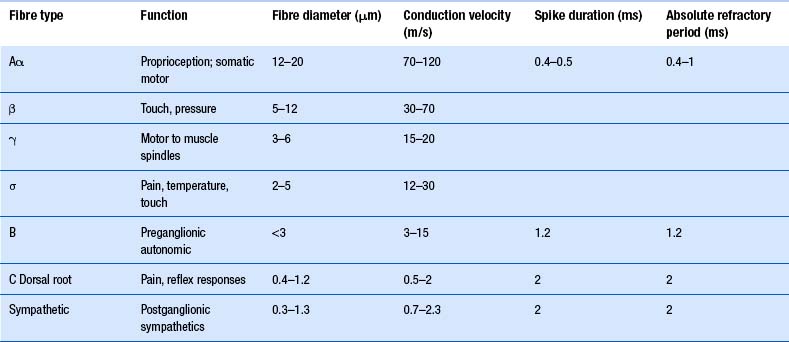
The axons of the various primary afferents display a variety of different diameters and myelination densities. Functionally, these morphological differences result in axons that transmit impulses at different rates of speed or conduction velocities. Table 5.1 outlines the various conduction velocities and diameters of different primary afferent axons.
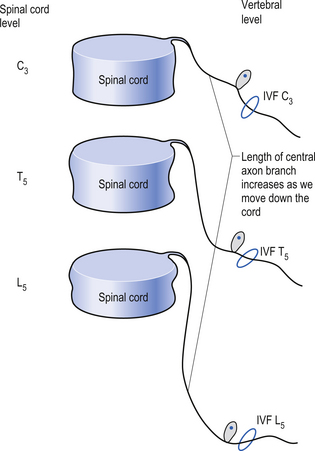
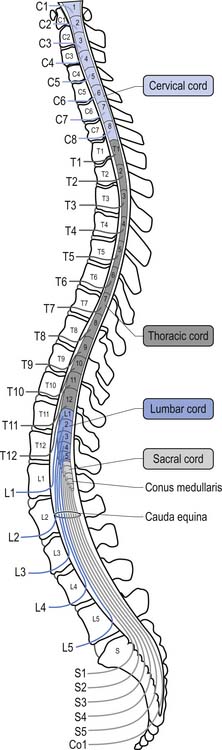
Because of different growth rate patterns between the spinal cord and the vertebral column the dorsal root ganglion cells below T3 are located at increasingly further distances from the segmental spinal cord levels that they supply. This results in the progressive lengthening of the central processes of the primary afferents and in the formation of the cauda equina after the spinal cord ends at the level of L1–L2 in most people. It is these central processes that may experience compression from a posterior central or posterior lateral vertebral disc herniation (Figs 5.2 and 5.3).
Receptors of primary afferent input
Structurally, receptors can be classified into three basic morphological forms: neuroepithelial receptors, visual receptors, and primary afferent receptors. In the case of the neuroepithelial receptor, the sensory sensitive detection apparatus is contained in the neuron cell body itself. These types of neurons are situated near the sensory surface with their axons projecting back towards the central nervous system. The unique quality of these receptors is that the neuron cell body has derived from epithelial tissue and remains in that tissue as a receptor organ. In humans the only known example of this type of sensory reception can be found in the sensory cells of the olfactory epithelium. Here, the axons of these neurons which form the olfactory nerve or 1st cranial nerve (CN I) traverse the cribriform plate of the ethmoid bone to synapse in the glomeruli of the olfactory bulb (Williams & Warwick 1984) (Fig. 5.4).
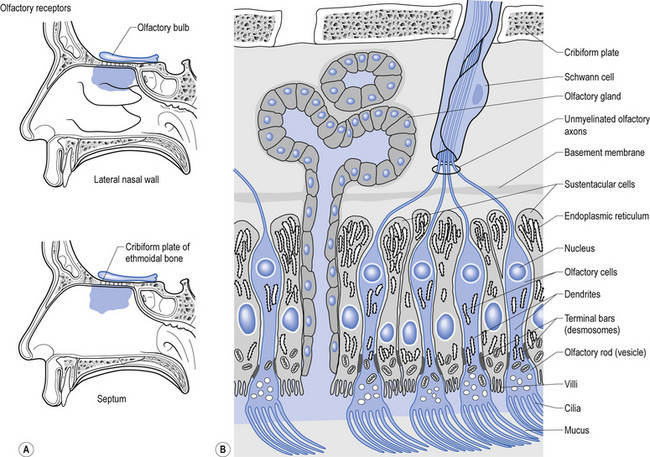
The second type of receptor can be classed as visual receptors. These receptors are similar to the neuroepithelial receptors in that the receptive apparatus is in their cell bodies; however, in this case, their cell bodies are derived from neuroectoderm of the fetal ventricles which form the fetal brain and migrate to the retinal areas (Fig. 5.5).
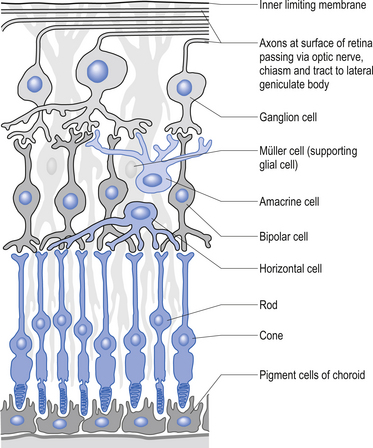
In the third type of receptor, which may be classified as a primary afferent receptor, the cell body is located near to or in the central nervous system. Long peripheral processes extend to the receptive area and form the specialised receptive structures that act as the receptors. All cutaneous receptors and most proprioceptors are composed of this type of receptor (Williams & Warwick 1984) (see Fig. 5.1).
 Quick facts 5.2
Quick facts 5.2The muscle spindle
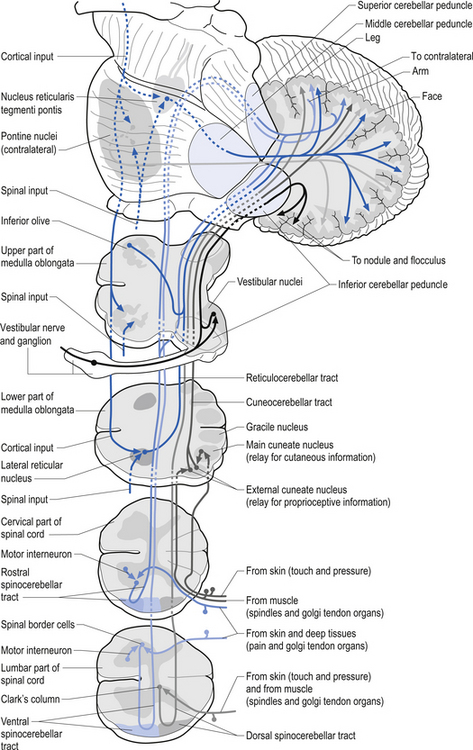
Muscle spindles provide the nervous system with information concerning the instantaneous static length of a muscle at any given moment and the rate of change of muscle length during any movement. The output of the muscle spindle is simultaneously transmitted to a variety of interneuron networks in the spinal cord. This information is then relayed to areas in the brainstem, cerebellum, and basal ganglia. The axons of the primary afferent neurons that relay muscle spindle information to the spinal cord form many collateral branches as they enter the spinal cord. Some of these collaterals proceed without synapsing on an interneuron to synapse directly (monosynaptic connection) on the alpha motor neuron pools of the muscle fibres from which they are relaying information (homonymous muscle), resulting in an excitatory stimulus (Fleshman et al. 1981). Other collaterals first synapse (polysynaptic connection) on interneurons in lamina VIII of the grey matter of the spinal cord, referred to as Clark’s column. These interneurons then project to the brainstem reticular formation nuclei, various cortical areas of the cerebellum via mossy fibres, and the basal ganglia (Cram & Kasman 1998). The spinocerebellar tracts are formed from the axons of these interneurons in Clark’s columns. The anterior spinocerebellar tracts receive axons from both ipsilateral and contralateral lamina VIII neurons. The posterior spinocerebellar tracts receive axons from only ipsilateral lamina VIII neurons. This is an example of reciprocal innervation that occurs when vital information transfer is required (Fig. 5.6). All regions other than the segmental interneurons and their associated alpha motor neurons receiving input from the primary afferents at that level are referred to as suprasegmental to the input. Most suprasegmental integration is involved with modulation of movement, or in feedback control of the accuracy of the movement generated. Some controversy exists as to whether the receptive input of muscle spindles can be consciously perceived. Since muscle spindles do not project directly to the somatosensory cortex, their activity was thought not to be consciously perceived. However, some research has indicated that conscious perception is possible (Williams & Warwick 1984).

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


