Chapter 11 Organization of the Brainstem
The spinal cord continues rostrally into the brainstem (Fig. 11-1), which performs spinal cord–like functions for the head. The brainstem contains the lower motor neurons for the muscles of the head and does the initial processing of general afferent information concerning the head. However, it does much more than this, reflecting in large part the additional functions of the cranial nerves attached to it, as well as some distinctive built-in brainstem functions.
The Brainstem Has Conduit, Cranial Nerve, and Integrative Functions
The cranial nerves contain not only the head’s equivalent of spinal nerve fibers but also those involved in the special senses of olfaction, sight, hearing, equilibrium, and gustation, or taste (Table 11-1). The olfactory and optic nerves project directly to the telencephalon and diencephalon, respectively, but the others project to or emerge from the brainstem. Thus a wide assortment of sensory and motor nuclei related to cranial nerve function can be found at various brainstem levels.
Table 11-1 Highly Simplified Overview of Cranial Nerve Functions*
| Cranial Nerve | Main Sensory Function | Main Motor Function |
|---|---|---|
| I | Smell | — |
| II | Sight | — |
| III | — | Eye movements, pupil and lens function |
| IV | — | Eye movements |
| V | Facial sensation | Chewing |
| VI | — | Eye movements |
| VII | Taste | Facial expression |
| VIII | Hearing, equilibrium | — |
| IX | Taste | Swallowing |
| X | Thoracic and abdominal viscera | Speech, swallowing; thoracic and abdominal viscera |
| XI | — | Head and shoulder movements |
| XII | — | Tongue movements |
* For more details, see Table 12-2 and Chapter 12 to 14.
It is clear that these three general types of activity are far from mutually exclusive. For example, ascending pathways to the thalamus arise not only in the spinal cord but also from cranial nerve nuclei; the latter therefore contribute to both conduit and cranial nerve functions. However, this parcellation does provide a useful framework on which to organize a treatment of the brainstem. It is difficult to learn about this portion of the nervous system all at once, so it is presented here in several parts. This chapter describes the overall anatomy of the brainstem and presents a series of sections showing the locations of some prominent nuclei and major ascending and descending tracts. The next three chapters describe the central connections of cranial nerves III to XII, and Chapter 15 presents a similar series of sections, labeled in more detail, with summary descriptions of the contents of various tracts and nuclei.
The Medulla, Pons, and Midbrain Have Characteristic Gross Anatomical Features
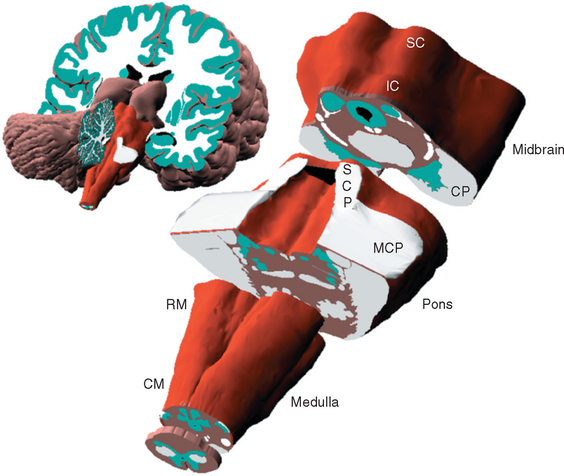
Each of the three major subdivisions of the brainstem—medulla, pons, and midbrain (Fig. 11-2)—has a characteristic set of surface features. As explained in subsequent sections, knowledge of these surface features can help make sense of the internal organization of the brainstem.
The Medulla Includes Pyramids, Olives, and Part of the Fourth Ventricle
The medulla is vaguely scoop shaped (Fig. 11-2). The “handle” corresponds to the caudal or closed portion, containing a central canal continuous with that of the spinal cord. The open portion of the scoop corresponds to the rostral or open medulla, in which the central canal expands into the fourth ventricle. The apex of the V-shaped caudal fourth ventricle, where it narrows into the central canal, is called the obex (Fig. 11-3A).
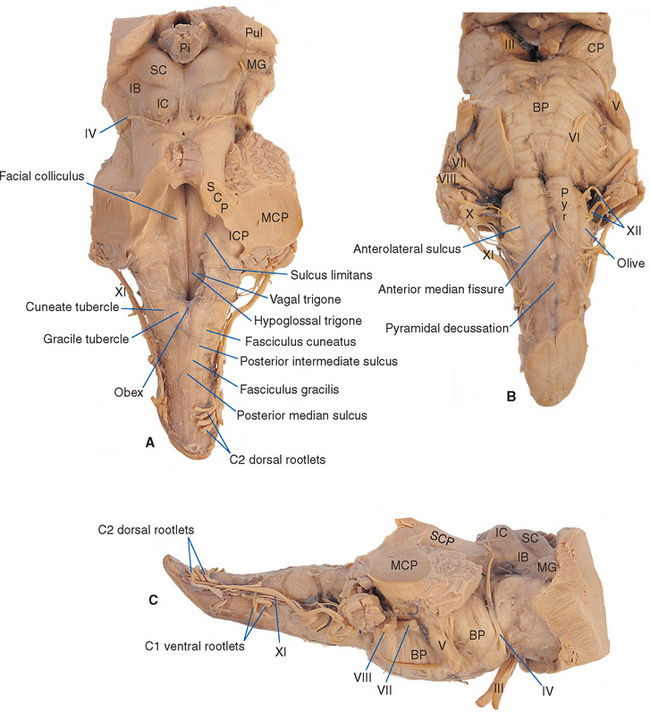
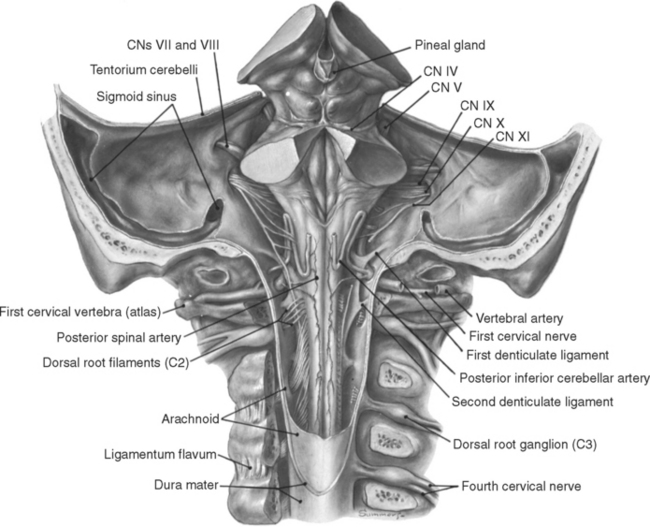
The longitudinal grooves on the surface of the spinal cord continue into the medulla, more clearly at some levels than at others. They divide the surface of the caudal medulla and part of the rostral medulla into a series of columns that completely encircle it (Fig. 11-3). The anterior median fissure is briefly interrupted by the pyramidal decussation at the junction between spinal cord and brainstem, but then it continues rostrally to the edge of the pons, separating the two pyramids (Fig. 11-3B). Proceeding around in a posterior direction, the anterolateral sulcus marks the other side of the pyramid. The rootlets of the hypoglossal nerve (XII) emerge from this sulcus, mainly in the rostral medulla. In the rostral medulla, the column dorsal to the hypoglossal rootlets is enlarged to form an oval swelling called the olive. The rootlets of the glossopharyngeal (IX) and vagus (X)* nerves emerge from a shallow lateral groove dorsal to the olive. The posterolateral sulcus also continues into the medulla; the area between it and the line of rootlets of cranial nerves IX and X (which in the spinal cord would overlie Lissauer’s tract and the posterior horn) overlies the spinal tract of the trigeminal nerve. As explained in the next chapter, the spinal tract of the trigeminal nerve is the head’s equivalent of Lissauer’s tract. Finally, the posterior columns continue into the medulla. Fasciculus cuneatus, adjacent to the posterolateral sulcus, extends rostrally to a small swelling called the cuneate tubercle, which marks the site of nucleus cuneatus. Fasciculus gracilis, adjacent to the midline, extends rostrally to a similar small swelling called the gracile tubercle, which marks the site of nucleus gracilis.
If the cerebellum is removed (as it has been in Fig. 11-3), one can peer down on the floor of the fourth ventricle. Here too, various grooves and elevations signify the presence of underlying nuclei. The sulcus limitans can often be followed rostrally along the floor of the ventricle into the pons (Fig. 11-3A). As in the embryonic spinal cord, it is a line of separation between motor nuclei (now medial to it) and sensory nuclei (now lateral to it; see Figs. 2-10 and 12-1). The portion of the medulla and pons in the floor of the ventricle, lateral to the sulcus limitans, is mostly occupied by vestibular nuclei and is referred to as the vestibular area. The area medial to the sulcus limitans overlies a series of motor nuclei, three of which make visible elevations. In the medulla, the hypoglossal nucleus and the dorsal motor nucleus of the vagus make small triangular swellings, appropriately called the hypoglossal and vagal trigones. Farther rostrally, in the pons, is another elevation called the facial colliculus. This elevation is not caused by an underlying motor nucleus of the facial nerve, as one might surmise from its name. Rather, it is the location of the abducens nucleus; fibers destined for the facial nerve loop over it at this location on their way out of the brainstem (see Figs. 12-6 and 12-7).
The Pons Includes the Basal Pons, Middle Cerebellar Peduncles, and Part of the Fourth Ventricle
The pons is dominated by the massive, transversely oriented structure on its ventral surface from which it derives its name (Figs. 11-3 and 11-4). Pons is the Latin word for “bridge,” and this portion of it (called the basal pons) looks like a bridge interconnecting the two cerebellar hemispheres. It is not, however, a direct interconnection. Rather, many of the fibers descending in each cerebral peduncle synapse in scattered nuclei of the ipsilateral half of the basal pons. These nuclei in turn project their fibers across the midline, after which they funnel into the middle cerebellar peduncle (brachium pontis*) and finally enter the cerebellum (see Fig. 11-12).
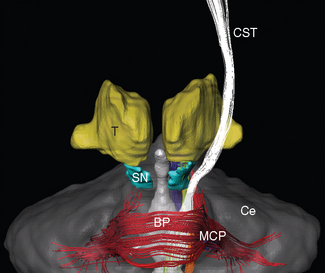
The trigeminal nerve (V) enters the brainstem at a midpontine level, and three others enter (or leave) along the groove between the basal pons and the medulla (Fig. 11-3). The abducens nerve (VI) is the smallest and most medially located of these three, exiting where the pyramid emerges from the basal pons. The facial nerve (VII) is farther lateral and consists of two parts: a larger and more medial motor root and a smaller sensory root (sometimes referred to as the intermediate nerve [see Fig. 3-17B]). The vestibulocochlear nerve (VIII) is slightly lateral to the facial nerve and also has two parts: a vestibular division and a more lateral cochlear division.
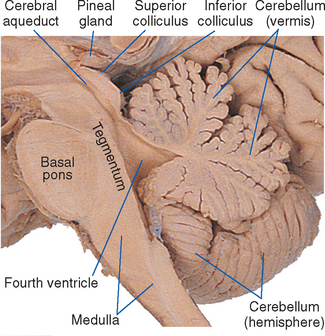
The superior cerebellar peduncle (brachium conjunctivum*) forms much of the roof of the fourth ventricle in the pons. It emerges from the cerebellum, moves toward the midline and the brainstem, and enters the latter near the junction between the pons and midbrain. At this same junction, the trochlear nerve (IV) emerges from the dorsal surface of the brainstem. The superior cerebellar peduncle is covered in the rostral pons by a flattened band of fibers called the lateral lemniscus, which forms part of the ascending auditory system and terminates in the inferior colliculus.
The Internal Structure of the Brainstem Reflects Surface Features and the Position of Long Tracts
At any given brainstem level rostral to the obex, three general areas can be identified in cross section (Fig. 11-5): (1) the area posterior to the ventricular space, (2) the area anterior to the ventricular space, and (3) large structures “appended” to the anterior surface of the brainstem. (In the caudal medulla the central canal is surrounded by structures, including some that are anterior to the ventricular space at more rostral levels.)
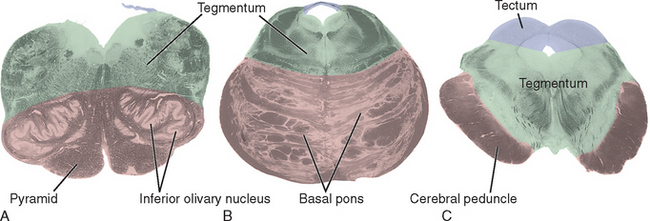
Although the brainstem is derived embryologically from a serial array of vesicles, in adults it no longer possesses an organization quite so neat (Fig. 11-4). For example, the basal pons usually extends rostrally to a point anterior to the tectum of the midbrain, and the pineal gland and part of the thalamus extend posteriorly so far that they overlap the tectum of the midbrain. Because the most instructive way to study the brainstem is to consider a series of parallel sections through it (all perpendicular to its long axis), I have ignored minor inconveniences such as the intrusion of the basal pons under the midbrain and have used, for purposes of the following discussion, reference transverse planes (Fig. 11-6) that subdivide the brainstem into six parts: caudal and rostral medulla, caudal and rostral pons, and caudal and rostral midbrain.
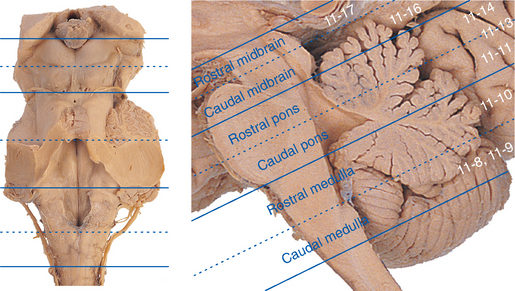
The following discussion points out major brainstem structures at these levels and the locations of tracts that begin or end in the spinal cord. The next three chapters deal with the cranial nerves and their tracts and nuclei. Finally, as noted previously, the material of all four chapters is integrated in a series of more extensively labeled sections in Chapter 15.
The Corticospinal and Spinothalamic Tracts Have Consistent Locations throughout the Brainstem*
The three major longitudinal pathways (corticospinal tract, posterior columns, and spinothalamic tract) that were followed through the spinal cord in Chapter 10 can be followed systematically through the brainstem, as indicated in Figure 11-7. Two of the three stay in more or less the same location throughout the brainstem. Corticospinal fibers travel in the most ventral part of the brainstem, traversing the cerebral peduncle, basal pons, and medullary pyramid. At the spinomedullary junction, most of the fibers in the pyramids decussate and form the lateral corticospinal tracts. The spinothalamic tract at all levels of the brainstem is in or near the anterolateral corner of the tegmentum, similar to its position in the spinal cord. The posterior columns terminate in the posterior column nuclei (nucleus gracilis and nucleus cuneatus) of the medulla. Efferent fibers from these nuclei decussate in the medulla to form the medial lemniscus, which reaches the thalamus. The medial lemniscus starts out near the midline and then moves progressively more laterally, rotating nearly 180 degrees as it proceeds through the brainstem.
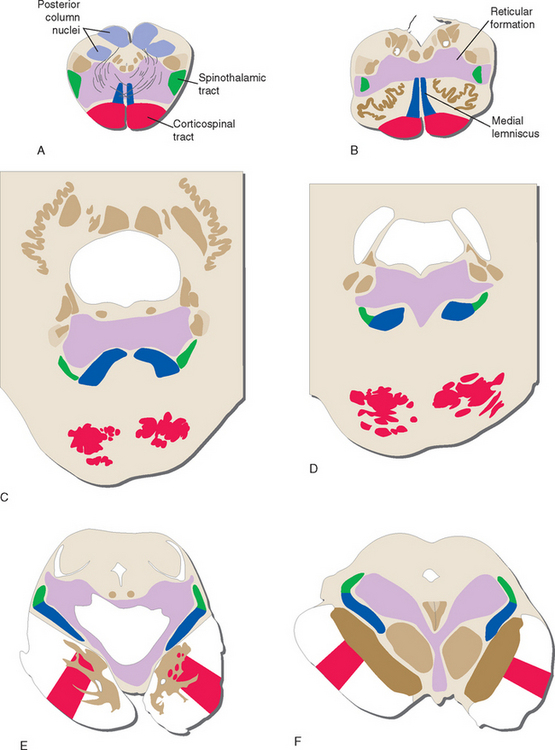
The Medial Lemniscus Forms in the Caudal Medulla
The caudal (closed) medulla extends from the caudal edge of the pyramidal decussation (where the medulla becomes continuous with the spinal cord) to the obex, which marks the caudal end of the fourth ventricle.
The caudal medulla (Figs. 11-8 and 11-9) looks somewhat like the spinal cord. Part of the anterior horn is still present caudally (Fig. 11-8), as are structures similar to Lissauer’s tract and part of the posterior horn. The latter two are actually the spinal tract and spinal nucleus of the trigeminal nerve. These are the head’s equivalent of Lissauer’s tract and the substantia gelatinosa (i.e., they deal with pain, temperature, and some tactile information, as discussed in Chapter 12).
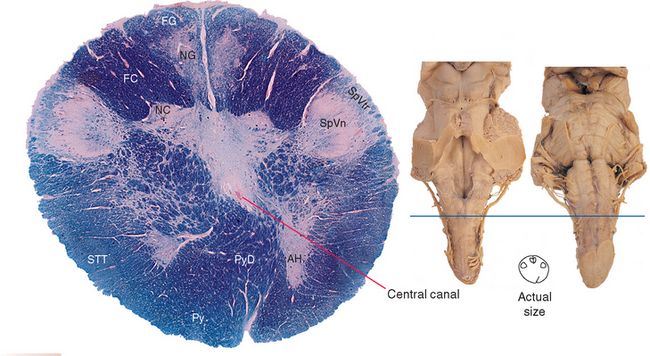
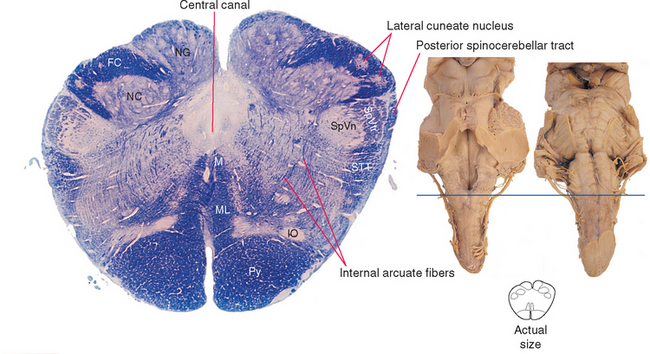
Fasciculi gracilis and cuneatus continue into the caudal medulla but are gradually replaced by the posterior column nuclei (nucleus gracilis and nucleus cuneatus). Nucleus cuneatus begins and ends a bit rostral to nucleus gracilis, so even in Figure 11-9, part of fasciculus cuneatus is still present. Postsynaptic fibers leave these two nuclei in a ventral direction and arch across the midline to form the contralateral medial lemniscus, a vertically oriented band of fibers (Fig. 11-9). These decussating fibers are part of the collection of internal arcuate fibers and are sometimes called the sensory decussation. Throughout the medulla, the medial lemniscus is organized so that fibers representing cervical segments are most posterior (i.e., as though the homunculus were standing upright in these cross sections).
Adjacent to nucleus cuneatus and embedded in fasciculus cuneatus is the lateral (or external) cuneate nucleus (Fig. 11-9). This is the upper extremity equivalent of Clarke’s nucleus, and the axons of these cells join the posterior spinocerebellar tract in the inferior cerebellar peduncle at a slightly more rostral level.
The prominent pyramids (Fig. 11-9) and their decussation (Fig. 11-8) are located most anteriorly in the caudal medulla. Each pyramid consists of corticospinal fibers that originated in ipsilateral cerebral cortex and are (mostly) bound for the contralateral anterior horn.
Most of the area traversed by internal arcuate fibers in Figure 11-9 is reticular formation. A casual observer looking at this region in photographs such as these would not see much. This is, to a first approximation, what distinguishes the reticular formation from the rest of the brainstem; the posterior column nuclei, for example, look like nuclei, whereas the reticular formation just looks like the uniform neural tissue filling the gaps between identifiable structures, forming a central core throughout the brainstem tegmentum (Fig. 11-7). In fact, however, the reticular formation is organized—but on a microscopic level, as discussed briefly later in this chapter.
The Rostral Medulla Contains the Inferior Olivary Nucleus and Part of the Fourth Ventricle
The rostral (open) medulla, as defined here, extends from the obex to the rostral wall of the lateral recess, where the inferior cerebellar peduncle turns posteriorly to enter the cerebellum. The rostral medulla (Fig. 11-10) no longer looks much like the spinal cord, partly because the walls of the embryonic neural tube have been pushed outward to form the floor of the fourth ventricle (see Fig. 2-10).
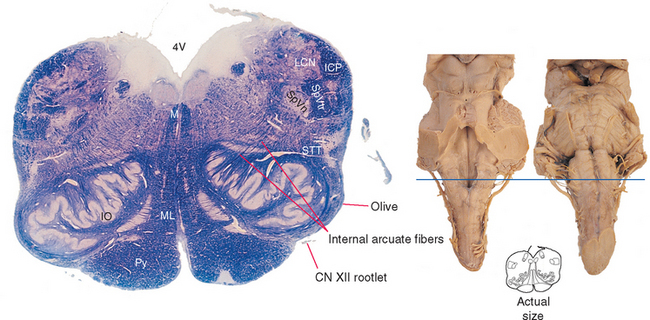
The caudal boundary (the obex) is approximately coincident with the caudal edge of the inferior olivary nucleus, a prominent structure that is responsible for the appearance of the olive as a surface swelling (Fig. 11-3). The inferior cerebellar peduncle is located dorsolaterally at these levels and grows progressively larger as it continues rostrally. Fibers can be seen leaving the medially facing mouth (or hilus) of the inferior olivary nucleus, arching across the midline, and joining the contralateral inferior cerebellar peduncle. These too are internal arcuate fibers. More and more are added at progressively more rostral levels of the medulla, increasing the size of the peduncle.
Medial to the inferior olivary nucleus is the medial lemniscus, which still has the shape of a flattened band with a dorsal-ventral axis. Anterior to the medial lemniscus is the pyramid. Fascicles of the hypoglossal (XII) nerve (Figs. 11-3 and 11-10) emerge lateral to the pyramid in the groove between it and the inferior olivary nucleus. Posterior to the medial lemniscus, near the floor of the fourth ventricle, is a small but distinctive bundle of fibers that can be followed all the way to the midbrain. This is the medial longitudinal fasciculus (MLF), which is involved in coordinating head and eye movements.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


