1 NORMAL NEURONAL CYTOLOGY AND STAINING The cytologic appearance of neurons varies depending upon their location. In general, neurons have moderate to abundant cytoplasm and a relatively large round nucleus with a prominent nucleolus. Projecting from the neuronal cell body are branching dendrites and a single axon. A range of tinctorial and immunohistochemical staining techniques are used for demonstrating neurons and their processes (Table 1.1). Table 1.1 Histologic demonstration of neurons: techniques and comments concerning their applications Conventional staining Hematoxylin and eosin Good for assessing general cytoarchitecture Nissl stain (e.g. cresyl fast violet) Good for assessing general cytoarchitecture. Allows estimation of cell density in thick sections; may be combined with stains for myelin (e.g. Luxol fast blue). Axon silver impregnation techniques Good for demonstration of axons and some neuronal inclusions. Used in conjunction with myelin stains to distinguish between demyelination and fiber degeneration Golgi stain Allows visualization of fine detail of neuronal cell processes. A technically difficult stain to perform which depends on block impregnation Immunohistochemistry Neurofilament proteins Strongly expressed in perikaryal and axonal cytoplasm (in normal neurons, perikaryal neurofilament proteins are non-phosphorylated and axonal neurofilament proteins phosphorylated) NeuN Neuron-specific nuclear protein, strongly expressed in neuronal nuclei. NeuN antibodies also show weak labeling of the neuronal cytoplasm Neuron specific enolase Strongly expressed in neuronal and axonal cytoplasm Synaptophysin Antibodies to synaptophysin detect neurosecretory vesicles, which are mainly located at synapses, with little staining of neuronal cell bodies PGP9.5 An antibody to neuron-specific ubiquitin C-terminal hydrolase, which is abundant in neurons and neuronal cell processes Chromogranin A Antibodies detect dense-core neurosecretory vesicles, which are sparsely distributed within the perikaryon of some neurons and concentrated at the synapses. Moderate staining of neuronal cell bodies Neurotransmitter-related Antibodies allow detection of neurotransmitter substance or enzyme involved in its biosynthesis Axonal degeneration inevitably follows the death of the neuron of which it is a part. A severed or severely damaged axon undergoes distal degeneration without usually provoking death of the proximal part of the neuron, although this does undergo a series of structural and metabolic changes (i.e. axon reaction and chromatolysis, see below). Within days, the distal part of the axon fragments and the surrounding myelin sheath breaks up into ovoids. Over the next 3 weeks or so, the axon and myelin debris are taken up by macrophages, which infiltrate the degenerating fiber tracts. Several methods can be used to demonstrate degenerating nerve fibers in the CNS. These include specialized silver impregnation techniques and stains for degenerating myelin and for lipid (Fig. 1.1). 1.1 Tract degeneration. Damage to the axon provokes a series of morphologic and biochemical changes in the neuronal cell body. These are collectively referred to as the axon reaction (Figs 1.2, 1.3). The changes include disruption and dispersion of Nissl bodies (chromatolysis) associated with rearrangement of the cytoskeleton and marked accumulation of intermediate filaments. The axon reaction is conspicuous in large neurons with axons that project into the peripheral nervous system and in some of the larger neurons with central projections. Chromatolysis is not visible on conventional light microscopy of small neurons or certain large neurons such as the cerebellar Purkinje cells, but changes can be demonstrated in these cells by electron microscopy and immunohistochemistry. 1.2 Axon reaction. Swollen or ballooned neurons are a feature both of the axon reaction and of a variety of diseases in which perikaryal changes occur independently of axonal damage (Table 1.2). Histologically, they appear as distended, weakly-staining cells with large, relatively clear nuclei (Fig. 1.4). Occasionally the cells contain small vacuoles. In conventionally processed tissues an artefactual lacuna is often present around the abnormal cell. Swollen or ballooned neurons can be demonstrated with several immunohistochemical markers (Fig. 1.5, Table 1.3). Table 1.2 Causes of ballooning of neurons Physiologic Anterior horn motor neurons, with age Nutritional Pellagra (niacin deficiency): spinal, brain stem, and cortical neurons Developmental Localized cortical dysplasia with cytomegaly Neurodegenerative Several types of frontotemporal lobar degeneration, including Pick’s disease and FTDP-17: neocortical and some basal neurons Prion disease Cortical and some basal neurons Table 1.3 Immunohistochemical profile of swollen neurons Neurofilament protein Accumulation of highly phosphorylated high-molecular-weight neurofilament protein, which is normally restricted to the axon αB-crystallin Accumulation of αB crystallin, which is not normally expressed by neurons Tau protein Accumulation of abnormally phosphorylated tau protein by a proportion of swollen neurons in corticobasal degeneration and Pick’s disease Ubiquitin Variably increased immunoreactivity for ubiquitin-protein conjugates PGP9.5 Variably increased immunoreactivity for this ubiquitin C-terminal hydrolase 1.4 Swollen neurons. 1.5 αB-crystallin immunoreactivity. Neurons within some nuclei in the CNS atrophy and degenerate in response to deafferentation. Examples are neurons in the lateral geniculate nucleus, which degenerate after optic nerve or tract lesions, and neurons in the pontine nuclei, which degenerate after interruption of descending frontopontine afferent fibers. Neurons in the inferior and accessory olivary nuclei undergo an unusual form of trans-synaptic degeneration after a destructive lesion (such as an infarct) of the ipsilateral central tegmental tract (Fig. 1.6). The olivary ribbon as a whole becomes thickened and neurons show marked enlargement, cytoplasmic vacuolation, and some dispersion of Nissl bodies. Olivary hypertrophy is associated with the development of palatal myoclonus in some patients. 1.6 Hypertrophy of the left olive due to a lesion involving the left central tegmental tract. Neurons are especially vulnerable to damage from hypoxia, which causes the following distinctive histologic changes (Fig. 1.7): 1.7 Hypoxic cell change in neurons. Neurons in certain parts of the brain that are especially vulnerable to hypoxic damage are: The pattern of regional susceptibility to hypoxia differs between infants and adults (see Chapters 2 and 8). Although dead neurons usually undergo liquefaction or are removed by phagocytosis, they may instead become encrusted and replaced by mineral salts, a process termed ferrugination. This is particularly prominent in the infant brain in response to hypoxic–ischemic damage (Fig. 1.8). 1.8 Ferruginated neurons. These are small spherical nuclear inclusions that are brightly eosinophilic and are often seen in neurons of the adult substantia nigra (Fig. 1.9a). They may also occur in other neurons, such as the pyramidal cells of the hippocampus and neurons in the tegmentum of the brain stem. Marinesco bodies are most common in the elderly and in dementia with Lewy bodies. They have an increased prevalence in neurons containing Lewy bodies. 1.9 Marinesco bodies. Ultrastructurally, Marinesco bodies are composed of filaments with the same diameter as intermediate filaments, and may be derived from the nuclear lamins. They are immunoreactive for ubiquitin, an 8 kD polypeptide involved in the degradation of many abnormal or short-lived proteins (Fig. 1.9b). CYTOSKELETAL AND FILAMENTOUS INCLUSIONS These are brightly eosinophilic rod-shaped or elliptical cytoplasmic inclusions that may appear to overlap the edge of a neuron (Fig. 1.11). They are immunoreactive for actin, actin-associated proteins, and caspase-cleaved TDP43. 1.11 Hirano bodies. These eosinophilic cytoplasmic inclusions occur in large neurons of the caudate nucleus. The inclusions resemble loose bundles of twigs and, like the thalamic neuronal inclusions described above, are common in patients with myotonic dystrophy. However, they are sometimes an incidental finding in elderly patients who do not have neurologic or muscular disease. The rod-like inclusions stain strongly with phosphotungstic acid/hematoxylin (Fig. 1.12). 1.12 Rod-like cytoplasmic inclusions in the caudate nucleus. There are several other types of neuronal inclusion that comprise or include elements of the cytoskeleton and usually occur in the context of specific neurodegenerative diseases (Table 1.4). These inclusions are described in more detail and illustrated in the sections concerned with the relevant diseases. Table 1.4 Examples of inclusion bodies in specific conditions and diseases These are composed of polyglucosans (polymers of sulfated polysaccharides) and are similar to corpora amylacea in composition and staining characteristics (see below). They are present in large numbers in Lafora’s disease (see Chapter 7), both in the CNS and in certain peripheral tissues such as sweat glands, liver, and skeletal muscle. Lafora body formation has been linked to aberrant glycogen hyperphosphorylation. The inclusions usually have a round core that is intensely periodic acid–Schiff (PAS)-positive (Fig. 1.13). Spicules of the core may radiate outwards, into a surrounding zone of less intensely PAS-positive material. A characteristic feature of aging is the development of ill-defined eosinophilic inclusions in the neurons of the inferior olivary nuclei. These inclusions are intensely immunoreactive with antibodies to ubiquitin (Fig. 1.14) and should not be confused with lipofuscin, which also accumulates with age (see below) and is particularly abundant in neurons of the inferior olivary nuclei. Colloid inclusions are round eosinophilic inclusions that usually occur in neurons in the hypoglossal nuclei (Fig. 1.15), but may be seen in other large neurons, particularly in the elderly. Electron microscopy shows that these inclusions result from dilatation of the endoplasmic reticulum by amorphous material. The importance of recognizing colloid inclusions, which do not have any clinical significance, is that they are occasionally confused with inclusions that are clinically significant such as Lewy bodies, pale bodies, and hyaline inclusions of motor neuron disease (see Chapter 27). These are small beaded eosinophilic inclusions seen in motor neurons in motor neuron disease. Cystatin C, transferrin, and peripherin have been detected in Bunina bodies. Ultrastructurally, they appear as electron-dense membrane-bound bodies (see Chapter 27). The acid vesicle system consists of endosomes, lysosomes, and lysosome-derived dense bodies. Lipofuscin (Fig. 1.16) is produced by oxidation of lipids and lipoproteins within the lysosomal system. It appears as orange-brown granular material in sections stained with hematoxylin and eosin, and is acid-fast (as demonstrated by the long Ziehl–Neelsen method) and autofluorescent under ultraviolet light. The granules are also sudanophilic and stain with PAS and Schmorl’s stain. Lipofuscin accumulates with aging in neurons and glia, particularly in: 1.16 Lipofuscin. Granulovacuolar degeneration. This term describes the accumulation of vacuoles containing small round dense bodies (granulovacuoles) (Fig. 1.17). The dense bodies are ubiquitinated and react with antibodies to some epitopes of the microtubule-associated tau protein. The immunohistochemical data have been interpreted as suggesting that dense bodies are derived from partial degradation of tau protein within lysosomes. Although anti-phosphoTDP43 antibody is sensitive for detecting granulovacuolar degeneration, TDP43 proteinopathies are not associated with an increase in granulovacuoles. Granulovacuolar degeneration is seen in normal aging after the sixth decade, predominantly in the hippocampal formation. Neurons in the CA1 field are most severely affected and, in descending order of severity, those in the prosubiculum, CA2, CA3, and CA4 fields. The density of hippocampal neurons showing granulovacuolar degeneration is increased in patients with Alzheimer disease and Pick’s disease, in whom granulovacuolar degeneration may also occur in neurons in the subcortical nuclei. 1.17 Granulovacuolar degeneration.
Pathologic reactions in the CNS
PATHOLOGIC RESPONSES IN NEURONS
ABNORMALITIES OF NEURONAL MORPHOLOGY
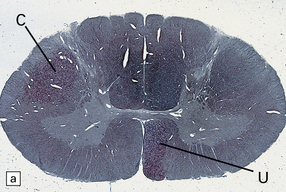
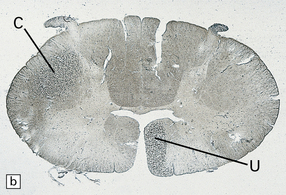
Adjacent sections of cervical spinal cord 1 month after an infarct involving the internal capsule. The infarct has caused degeneration of the descending crossed (C) and uncrossed (U) corticospinal tract fibers. (a) Stained with Oil red O. (b) Stained with Marchi’s method.
AXON REACTION AND CHROMATOLYSIS
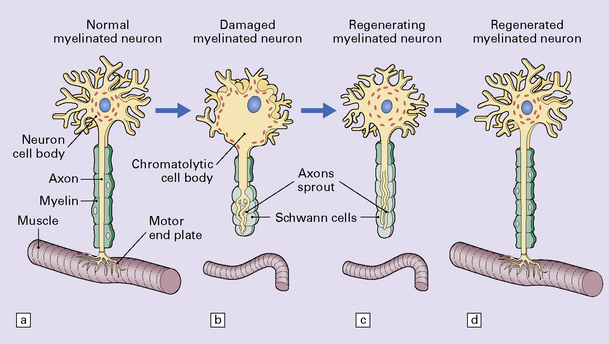
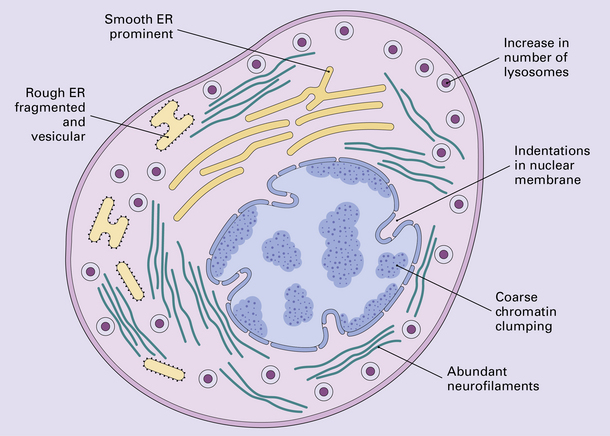
The changes that constitute the axon reaction have been most extensively investigated in anterior horn cells after peripheral nerve injury, as depicted in this figure, but neurons undergo similar alterations after injury to axons in the CNS. (a) A normal neuronal cell body is characterized by well-organized dendritic branches making synaptic contact with other neurons. Nissl bodies are prominent. (b) Following axonal damage there is degeneration of the peripheral axon. Dendrites that normally form synapses retract and synaptic contacts are lost. The rough endoplasmic reticulum becomes reorganized into small vesicular elements. The neuron inactivates genes coding for high molecular weight neurofilament protein (NFH) and switches on genes for peripherin. Neurofilaments and peripherin accumulate in the neuronal cell body. At this stage the neuronal cell body is swollen, shows only weak acidophilia, and lacks Nissl bodies. Denervated muscle fibers atrophy. (c) The axon regenerates in the peripheral nervous system (but not within the CNS). NFH synthesis is re-established with axonal elongation. (d) If the target tissue is reinnervated, the neuronal dendrites re-establish synaptic contact with other neurons and the normal cytologic appearance of the cell body is regained.
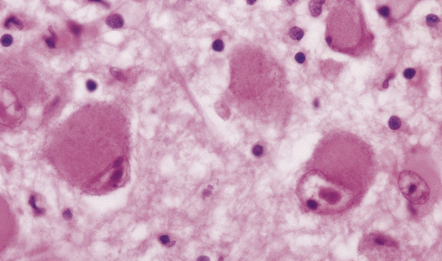
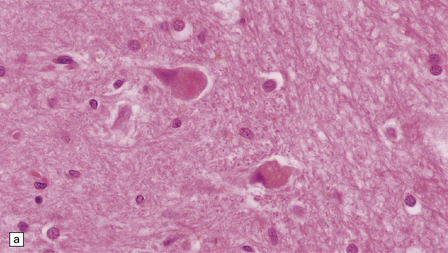
SWOLLEN NEURONS
Cortical dysplasia with hemimegalencephaly
Tuberous sclerosis
Corticobasal degeneration: neocortical and some basal neurons
Swollen neurons can also occur in Alzheimer disease, argyrophilic grain disease, progressive supranuclear palsy and several other neurodegenerative diseases
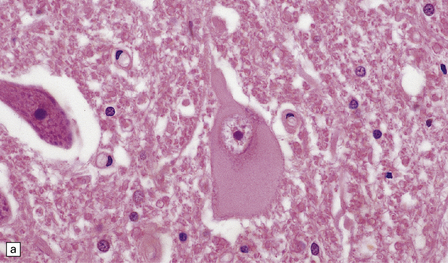
The cytoplasm lacks Nissl bodies and is weakly eosinophilic. The nucleus is large and vesicular. (a) Example from the anterior horn of the spinal cord. (b) Example from the cerebral cortex in corticobasal degeneration (see Chapter 28).
This is a characteristic feature of swollen neurons in most conditions. The protein is not, however, expressed by swollen neurons in pellagra (see Chapter 21), suggesting that neuronal swelling in this disorder has a different mechanism.
TRANS-SYNAPTIC NEURONAL DEGENERATION AND OLIVARY HYPERTROPHY
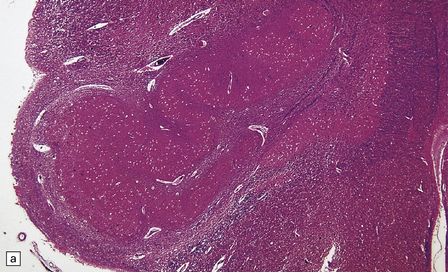
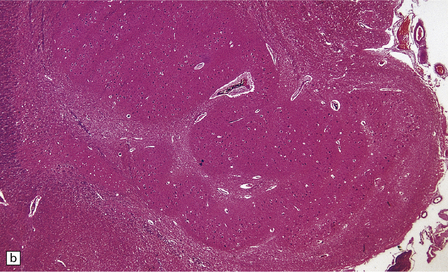
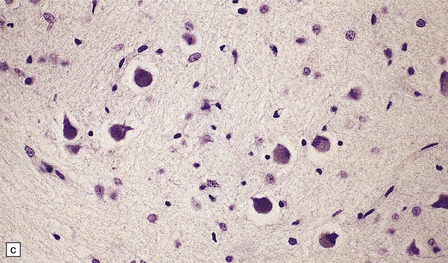
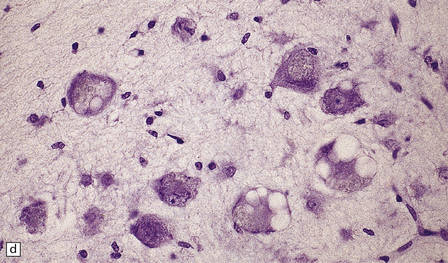
(a) Normal right olive. (b) Hypertrophy of the left olive. The lesion involving the left central tegmental tract is not shown. Compare the normal-sized neurons in the right olive with the enlarged vacuolated neurons in the left olive. (c) Normal neurons of the right inferior olivary nucleus stained with cresyl violet. (d) Neurons of the hypertrophic left inferior olive are enlarged and vacuolated.
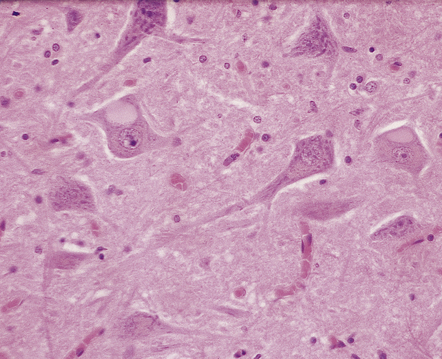
HYPOXIC CELL CHANGE
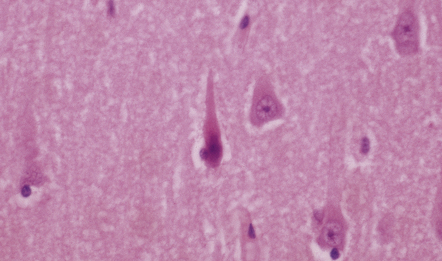
Affected neurons become shrunken and eosinophilic. The nuclei become condensed and lose their crisp contours. In this illustration the neuron in the center is affected while surrounding neurons appear normal.
 Microvacuolation (visible only at an ultrastructural level), due to swelling of endoplasmic reticulum and mitochondria.
Microvacuolation (visible only at an ultrastructural level), due to swelling of endoplasmic reticulum and mitochondria.
 Shrinkage of the cell body and increasing cytoplasmic acidophilia.
Shrinkage of the cell body and increasing cytoplasmic acidophilia.
 Condensation of nuclear chromatin and nuclear pyknosis.
Condensation of nuclear chromatin and nuclear pyknosis.
 pyramidal neurons in the CA1 field of the hippocampus
pyramidal neurons in the CA1 field of the hippocampus
 pyramidal neurons in layers 3 and 5 of the neocortex
pyramidal neurons in layers 3 and 5 of the neocortex
FERRUGINATION
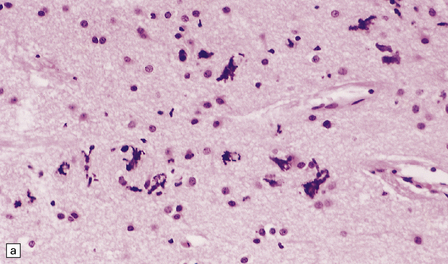
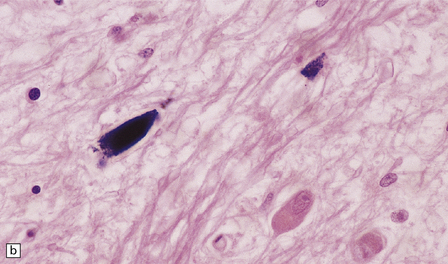
Ferruginated neurons become encrusted in mineral deposits and appear dark purple in sections stained with hematoxylin and eosin. The mineral deposits contain both calcium and iron and can be stained by Perls’ method and von Kossa’s technique. (a) Many ferruginated neurons in the cortex of a neonate who had hypoxic brain damage. Astroglial cells are not affected. (b) Higher power illustration of neurons replaced by mineral deposits.
NUCLEAR INCLUSIONS
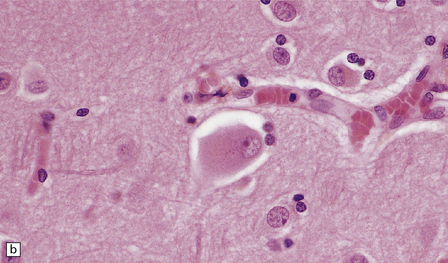
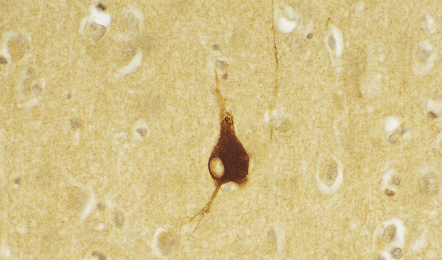
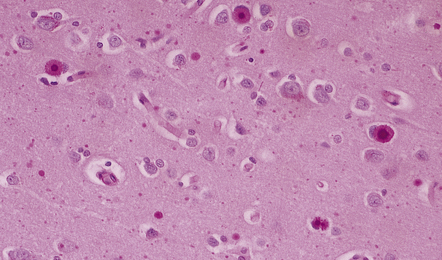
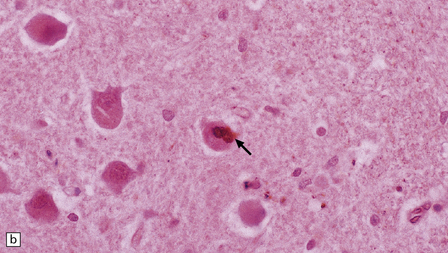
MARINESCO BODIES
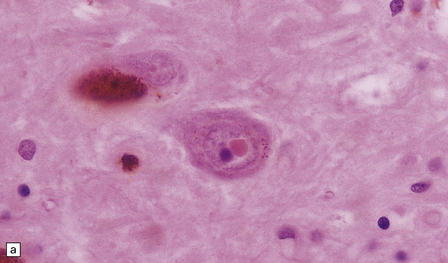
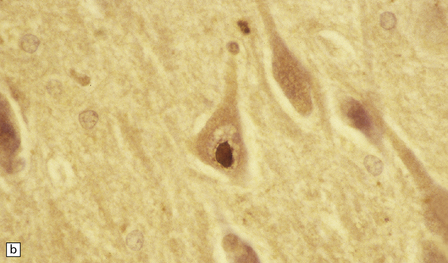
These intranuclear inclusions are visible in several neuronal groups. (a) Marinesco bodies are brightly eosinophilic intranuclear inclusions about the same size as a large nucleolus. (b) As shown here, they are strongly immunoreactive for ubiquitin.
NEURONAL CYTOPLASMIC INCLUSIONS
Hirano bodies
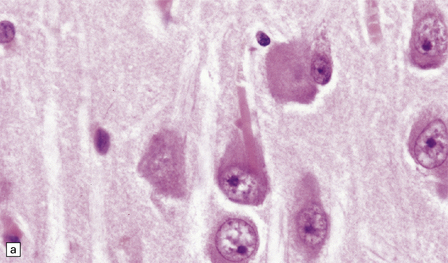
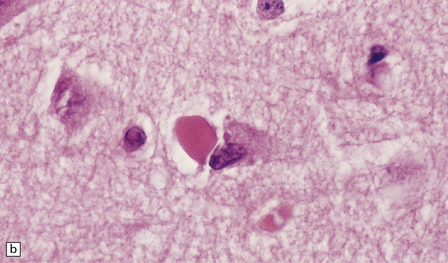
These are brightly eosinophilic inclusions of varied shape. (a) The majority are small rod-shaped structures with rounded ends. (b) Others have a globular appearance.
ROD-LIKE CYTOPLASMIC INCLUSIONS
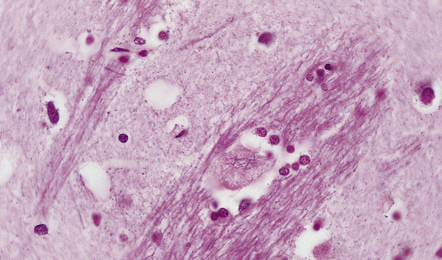
These occur exclusively in large neurons in the caudate nucleus. The inclusions may be an incidental finding, particularly in the elderly, but are most common in myotonic dystrophy. (Courtesy of Dr D A Hilton, Derriford Hospital, Plymouth.)
OTHER FILAMENTOUS NEURONAL INCLUSIONS
Inclusion
Association
Main constituents
Lewy bodies
Aging, Parkinson’s disease, dementia with Lewy bodies
α-synuclein, neurofilament protein, and ubiquitin
Neurofibrillary tangles
Aging, Alzheimer disease, progressive supranuclear palsy, post-encephalitic parkinsonism, Guam parkinsonism–dementia, myotonic dystrophy, subacute sclerosing panencephalitis, Niemann–Pick disease type C, other rare disorders
Phosphorylated tau protein (3R, 4R), ubiquitin
Pick bodies
Pick’s disease
Neurofilament protein, phosphorylated tau protein (3R), ubiquitin
MND inclusions
Motor neuron disease/amyotrophic lateral sclerosis
TAR DNA-binding protein 43 (sporadic cases) or superoxide dismutase 1 (some familial cases) or fused-in-sarcoma protein (other familial cases) ubiquitin, p62
CYTOSOLIC INCLUSIONS
EOSINOPHILIC INCLUSIONS IN THE INFERIOR OLIVES
MEMBRANE-BOUND CYTOPLASMIC INCLUSIONS
BUNINA BODIES
INCLUSIONS DERIVED FROM THE ACID VESICLE SYSTEM
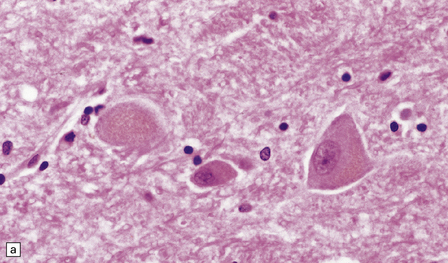
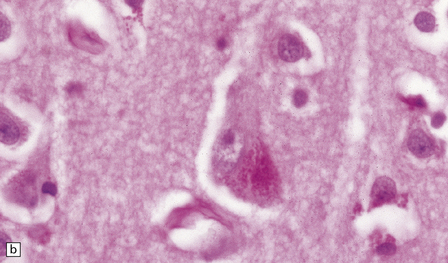
This accumulates with age in neurons and glia. (a) It has a golden brown color in sections stained with hematoxylin and eosin. (b) Lipofuscin stains strongly with PAS.
 Pyramidal neurons in the cerebral cortex and hippocampus.
Pyramidal neurons in the cerebral cortex and hippocampus.
 Large neurons in the amygdala, thalamus, and hypothalamus.
Large neurons in the amygdala, thalamus, and hypothalamus.
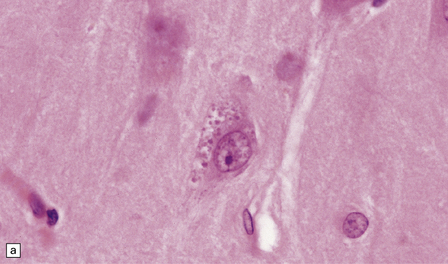
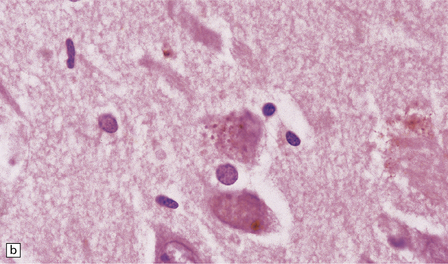
(a) These are small round basophilic bodies within otherwise clear vacuoles. They are mainly found in pyramidal neurons of the hippocampus. (b) The granules are variably immunoreactive for ubiquitin.![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Pathologic reactions in the CNS





Only gold members can continue reading. Log In or Register to continue


