Chapter 137 Sex Differences and Menstrual-Related Changes in Sleep and Circadian Rhythms
Abstract
Sex Differences in Sleep from Infancy to Adulthood
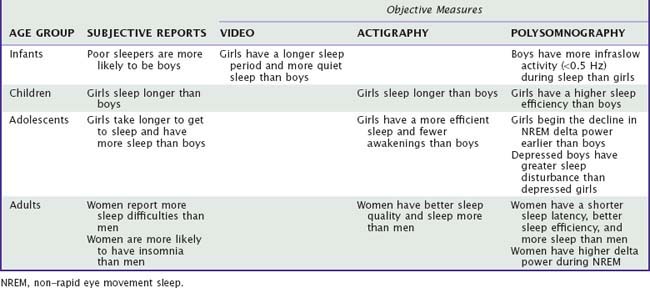
Sexual dimorphism describes morphologic differences between the sexes, although it also describes any biological process that differs between men and women. Extensive evidence from fruit flies to humans demonstrates sexual dimorphism in structure, function, and regulation of the brain. Differences between male and female brains are believed to result from the actions of gonadal secretions during a critical period of brain development. Data from animal models show that androgens produced by the testes in fetal and neonatal life induce sex differences in the neural structure and function of the brain, and such sex differences are also apparent in humans. Gonadal secretion is not the only mechanism: Gene expression in neuronal cells also plays a role in sexual dimorphism of the brain before gonadal secretion occurs. There is provocative evidence that even mechanisms of cell death in fetal or newborn brains are not identical between male and female infants.1 Other examples of sexual dimorphism include circadian clock genes, respiratory control, stress responses and hypothalamic-pituitary-adrenal axis, density of hypothalamic nuclei and sex hormone receptors in the suprachiasmatic nucleus (SCN), and the action of sex and reproductive hormones on sleep-regulatory mechanisms, all of which have implications for sleep and circadian rhythm regulation. Despite this, research into sex differences in sleep and circadian rhythm regulation lags behind other studies of sex differences in both humans and animal models. The following sections highlight current literature in sleep and circadian rhythms from infancy to adulthood, focusing on differences between the sexes. Major findings are summarized in Table 137-1.
Infancy
The development of sleep states and circadian rhythms in neonates has been reported in numerous studies, although few have reported on comparisons between the sexes. Of those that have compared male and female infants, findings are contradictory. Some studies using nighttime video recordings of infants find no differences in organization of sleep–wake state,2 but others find that female infants have more mature proportions of quiet sleep and longer sleep periods than male infants have.3 However, based on electroencephalograph (EEG) recordings in healthy infants during the first few months of life, no sex differences in sleep states were found,4 although newborn boys have more infraslow activity (<0.5 Hz) during sleep than newborn girls have,5 supporting the hypothesis that the central nervous system (CNS) of newborn girls is more mature than that of boys. Female infants also have more mature respiration than male infants during the first 6 months of life—possibly related to accelerated CNS maturation.6
In healthy infants, the ratio of nighttime sleep to total sleep becomes progressively greater with age and demonstrates a more-organized pattern of sleep, particularly in female infants. Further, the time course of REM sleep development corresponds with brain maturation,7 and higher proportions of REM sleep, together with a maturational lag in REM sleep, have been reported in infants at risk for sudden infant death syndrome (SIDS),8 a syndrome in which there is a clear male predominance.
The evidence linking mutations of circadian clock genes and sleep- and circadian-related disorders is compelling.9 Further, sleep disturbances are important symptoms of many psychiatric disorders. There is increasing evidence that delayed development of circadian rhythms in infancy contributes to differential risk for disease, particularly psychiatric disorders, between male and female infants, as reviewed in the following section.
Childhood
Survey Data
Worldwide, several cross-sectional survey-based studies of hundreds of schoolchildren have found that girls report sleeping longer than boys.10,11 However, other large, survey-based, longitudinal studies have failed to find any sex differences in sleep parameters.12 Lack of consistency between findings might result from factors such as cultural or racial differences and the limitations of using questionnaires to investigate sleep.
Actigraphy Data
Data from actigraph recordings have been more consistent in regard to sex differences in sleep.13,14 Several studies have found that girls sleep more and spend more time in motionless sleep than boys. Minority boys tend to report shorter sleep times than white boys.15 Such findings raise interesting questions about the role of racial background in sleep.
Polysomnographic Data
Although many reports of sleep in children include polysomnographic (PSG) data, few studies report on the relation of sleep to sex. However, significant sex-by-age-group differences on several PSG measures have been reported in children aged 3 to 8 years.16 Younger girls (ages 3 to 5 years) appear to have higher sleep efficiency, smaller proportion of wake, and a higher proportion of stage 3 sleep than boys of similar age. Among older children (≥6 years), girls appear to have a higher proportion of stage 1 sleep than boys. Despite these significant findings, the effect sizes for sex differences are quite small, and therefore their clinical significance is unclear. Based on measures of the cyclic alternating pattern, which refers to EEG activity characterized by sequences of transient episodes that differ from the background of basal EEG, sex differences have been found, suggesting that girls have greater NREM instability, even when standard EEG criteria showed no sex differences.17 However, another study using similar methodology did not find these sex differences.18
Additional studies relevant to sex differences in children’s sleep include those differentiating healthy children from children with medical problems such as psychiatric disorders. Significant age-related differences in sleep macroarchitecture that vary by sex and disease have been reported.19 Depressed adolescent boys have the most sleep disturbance compared to nondepressed boys, nondepressed girls, and depressed girls. However, differences in sleep macroarchitecture are minimal between depressed and nondepressed girls.19 Pubertal boys and girls in the depressed and nondepressed groups slept significantly less than their prepubertal counterparts. Early-onset depression is associated with a reduction in synchronization of sleep EEG rhythms (temporal coherence) that is stronger in depressed girls than boys.20 Girls with major depressive disorder (MDD) also demonstrate dampened amplitude of circadian rest–activity cycles, even in those who have yet to reach developmental maturity.21 By contrast, dampened circadian amplitude is not evident in depressed boys until adolescence. Together, these findings suggest that there may be a differential developmental influence on sleep in early depression that is dependent on sex.19
Girls at high risk for depression, based on maternal history, but who have yet to experience depression themselves may also show impaired synchronization of sleep EEG rhythms and reduced amplitude of circadian rest–activity cycles.22,23 This finding raises the possibility that it is the initial entrainment of circadian rhythms that is compromised in those with MDD and that this biological rhythm disturbance confers a risk for the future development of MDD. Preliminary evidence from the University of Michigan indicates greater sleep disturbance and slower entrainment of rest–activity cycles in infants from a maternal history of depression.24
Adolescence
Survey Data
Adolescence is a period of many changes in sleep patterns, which are under the influence not only of age but also of developmental or pubertal status, as well as changes in sleep schedule. Using questionnaire-based methodologies, several studies have found sex differences in sleep schedules; for example, girls can take longer to get to sleep than boys, boys might sleep longer than girls, or, conversely, boys might have slightly less sleep time than girls, even though they go to bed later and get up later than girls.25 Investigations of insomnia and pubertal development suggest that boys might have earlier onset of insomnia than girls and that onset of menses may be associated with an increased risk for insomnia.26 This increased risk for insomnia in girls after menarche corresponds with an increased risk for depression,27 which is a risk factor for insomnia. Clearly the relationship among insomnia, depression, and pubertal development is complex because several social and biological changes occur around the onset of menses, which can increase risk for both depression and insomnia.
Actigraphy Data
In studies using actigraphy, adolescent boys have been found to sleep less, have less-efficient sleep, and awaken earlier than girls, likely related to greater activity levels during the night in boys.28 Measures of total sleep time by actigraphy and PSG are in close agreement for adolescent girls, but actigraphy can underestimate total sleep time in boys, possibly because of the differential pattern of nocturnal movement between girls and boys.
Polysomnographic Data
In adolescence, a well-described EEG change in sleep is a steep decline in NREM slow-wave (delta) activity (SWA), and data from cross-sectional studies indicate that delta activity declines by about 50% between the ages of 10 and 20 years.29,30 Longitudinal data from cohorts of 9- and 12-year-olds followed up for 4 years suggest that girls begin the steep adolescent decline in SWA earlier than boys, which has been hypothesized to reflect an earlier onset of adolescent synaptic pruning in girls.31 It has long been speculated that sexual maturation and the decline in delta EEG activity in NREM sleep may be causally related. The decline in visually scored slow-wave sleep (SWS) is significantly correlated with increasing Tanner stage, a relationship that is independent of age.32 However, data collected longitudinally has shown that the decline in SWA during NREM sleep in boys and girls between the ages of 12 and 14 years is associated with age but not with pubertal stage.33 Feinberg34 has suggested that the delta decline during adolescence is a component of widespread brain reorganization, of which other manifestations include reductions in brain metabolic rate, decreased plasticity, and the emergence of adult cognitive capacity. It is possible that abnormalities in these maturational brain processes give rise to the emergence of abnormal circuits that cause psychiatric illnesses.
In children and adolescents with MDD, significant group-by-age-by-sex interactions for several sleep variables are observed.19 Depressed adolescent boys have the greatest sleep disturbance, and depressed girls do not differ significantly from healthy girls on sleep measures. However, temporal coherence of sleep EEG rhythms (a measure of synchronization of EEG frequencies of the same periodicity), are significantly lower in adolescents with MDD compared to healthy controls, and both sex and age can strongly influence the between-group differences. The lowest temporal coherence may be found among girls with MDD, even in those younger than 13 years.20 Strikingly, almost half of adolescents identified as having the most abnormal coherence values either showed symptoms of depression or met diagnostic criteria within 2 years.23 Low temporal coherence represents a more chaotic organization of sleep EEG rhythms, suggesting that early-onset depression is associated with a reduction in synchronization of sleep EEG rhythms that shows a differential maturational course in boys and girls.20 Further, low temporal coherence is evident in adolescent girls at high risk for, but not yet fulfilling criteria for, depression.
Adulthood
Survey Data
Sex differences in sleep–wake patterns in adults are commonly reported. Men are more likely to nap, and they report more disturbed, lighter sleep than women, particularly in older adulthood.35 However, women have an increased risk for insomnia across different age groups. In 1990 a meta-analysis was conducted on 27 studies addressing sex differences on 31 indices of sleep behavior of adults aged at least 58 years.36 Although no differences in total sleep time were evident, older men tended to show more changes from the patterns of healthy youthful sleep, with small to moderate effect sizes. Such differences included more time in bed, less delta sleep, and shorter REM latency for men.
Actigraphy Data
Actigraphic assessment of sleep has shown that women exhibit better sleep quality than men,37 with higher sleep efficiency index and lower frequency of transitions between sleep and wakefulness. Women tend to sleep more than men and have shorter sleep-onset latency. Regardless of sex, age affects sleep fragmentation because adults have a strong decline in actual sleep time and sleep efficiency, as well as increased sleep latency as they age.
Polysomnographic Data
In parallel to actigraphy, PSG data demonstrate better sleep quality in women than men, with shorter sleep onset latency and better sleep efficiency and more time asleep and less time awake. Women have higher delta power relative to men, although there is not a significant age-by-sex interaction,38 suggesting that the process of aging has a similar impact on delta EEG in men and women.
It has been argued that sex differences in sleep regulation may be subtle in healthy young adults but that they may become more obvious when the system is challenged, such as in instances of sleep deprivation, stressors, or in diseases associated with immune or hypothalamic-pituitary-adrenal axis abnormalities, such as MDD.39 For example, compared to young men, young women have a greater delta response to the stress of sleep deprivation,40 and sex differences in sleep normally seen in healthy adults become even more magnified when comparing depressed men to depressed women. Men with MDD show lower SWA than women with MDD or men and women without MDD, whereas depressed women show lower temporal coherence than controls.39 These disturbances in sleep microarchitecture that are more apparent in women with MDD might contribute to the increased risk of MDD observed in women.
There may be greater biological flexibility and adaptation among women, reflected as a greater response to a challenge condition. However, it has been hypothesized that women should be less responsive to a challenge in order to preserve sleep, neuroendocrine function, and circadian phase and amplitude.41 Indeed, a review reported that in general, between puberty and menopause, hypothalamic-pituitary-adrenal axis and autonomic responses tend to be lower in women compared to men of same age.42
The Menstrual Cycle and Effects of Ovarian Hormones on Sleep and Circadian Rhythms
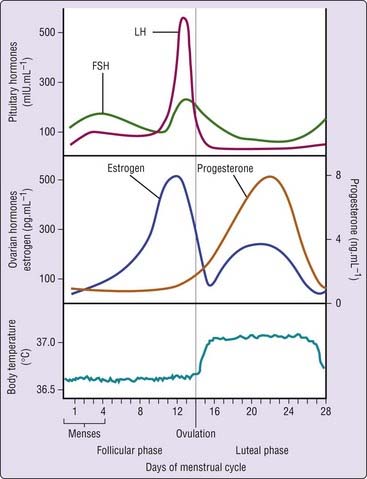
Conventionally, in a normal menstrual cycle of 28 days, day 1 is identified as the first day of bleeding (menses). Ovulation occurs around day 14, dividing the cycle into two phases: a preovulatory follicular phase and a postovulatory luteal phase (Fig. 137-1).43 Under the control of the hypothalamic–anterior pituitary axis, ovarian follicles grow during the follicular phase, with an associated rise in plasma estradiol. Circulating estradiol levels peak just before ovulation, triggering a surge in secretion of luteinizing hormone (LH) from the anterior pituitary. Ovulation occurs 12 to 16 hours later, around day 14. In the luteal phase, progesterone dominates, being secreted from the ruptured dominant follicle (corpus luteum), together with estradiol. Approximately 14 days after ovulation, if there is no implantation of a fertilized ovum, hormone levels drop precipitously, heralding the onset of menstruation. Ovulatory cycles are typically between 25 and 35 days. Most negative menstrual symptoms are experienced by women during the last few days of the cycle, as progesterone and estrogen levels decline, and during the first few days of menstruation.44

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


