Chapter 13 The Chemical Senses of Taste and Smell
Dating back to their origins in some primordial sea, living cells have shared an ability to respond to chemicals, at a minimum detecting and absorbing nutrients. Neurons detect chemicals at synapses, but some cells in or closely associated with the nervous system go beyond this, specializing in the detection of certain classes of chemicals adjacent to their membranes and using this information to affect autonomic function, behavior, or perception. These cells fall into four general categories*: (1) the myriad visceral chemoreceptors that work in the background, mostly inaccessible to conscious awareness, keeping track of the concentrations of oxygen, glucose, neuroactive hormones, and other substances; (2) gustatory receptor cells, or taste receptor cells, that mediate the sense of taste; (3) olfactory receptor neurons that mediate the sense of smell; and (4) chemosensitive endings, such as some trigeminal endings in mucous membranes, that mediate what has been termed the common chemical sense—sensations such as the heat of chili peppers, the sting of ammonia, or the coolness of menthol. The visceral chemoreceptors monitor internal chemical composition, and the other three monitor external chemistry, whether of local air or of substances being considered for ingestion.
Taste, smell, and the common chemical sense, like all the other senses of which we are consciously aware, have both rewarding and warning functions. In this case, the chemical senses are the basis not only of the enjoyment of a meal or a glass of wine but also of the detection of things such as spoiled food or smoke from a fire.
The Perception of Flavor Involves Gustatory, Olfactory, Trigeminal, and Other Inputs
The term taste as used in this book is synonymous with the gustatory sense, the set of sensations engendered by stimulating taste buds. We commonly assume that this is the same thing as the total sensation we perceive when eating or drinking, but such integrated sensations of flavor are actually the result of the combination of at least three different kinds of input: (1) direct chemical stimulation of taste buds, (2) stimulation of olfactory receptors by vapors from food (see Fig. 13-8), and (3) stimulation of chemical-sensitive and somatosensory free nerve endings of the trigeminal and other nerves in the mucous membranes of the oral and nasal cavities. The latter sets of endings respond to qualities such as the pungency, spiciness, temperature, and texture of food. We have difficulty appreciating the subtleties of foods and beverages using just our taste buds, and people with deficits in their sense of olfaction complain that things “taste” bland.
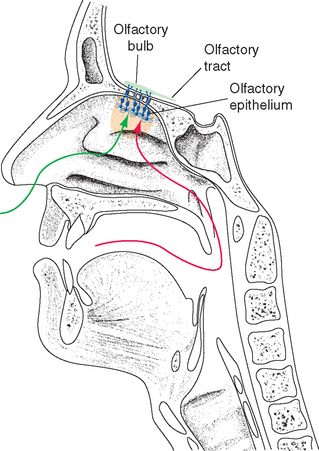
Although gustation, olfaction, and the common chemical sense are emphasized in this chapter, the full perception of flavor with all its nuances requires the integration of multiple inputs. Somatosensory aspects of flavor perception, for example, are more important than we usually realize. Some things “taste” better hot; others taste better cold. Some foods are designed to be chewy (e.g., some candies); others are expected to be tender or crunchy.* Vision also comes into play; the appearance of food affects not just our anticipation of it but also its palatability. Even the visceral chemoreceptors of which we are usually unaware come into play, as in the common experience of something tasting much better to a hungry eater. The orbital cortex of the frontal lobe is a major site where these multiple factors are combined (see Fig. 13-21).
Taste Is Mediated by Receptors in Taste Buds Innervated by Cranial Nerves VII, IX, and X
The Tongue Is Covered by a Series of Papillae, Some of Which Contain Taste Buds
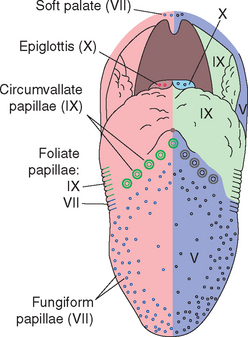
The tongue is mostly muscle, but its surface is covered by a series of bumps and folds called papillae (Fig. 13-1). Most are small conical projections not involved in taste, but fungiform, foliate, and circumvallate papillae contain taste buds (Fig. 13-2). About 200 to 300 fungiform (“mushroom-shaped”) papillae are scattered across the surface of the anterior two thirds of the tongue, concentrated on the tip and sides. Typical fungiform papillae contain 3 to 5 taste buds. Foliate (“leaflike”) papillae are the most posterior of a series of about 20 folds on the sides of the posterior tongue; each has 100 to 150 taste buds in its walls. Finally, a series of 8 to 9 circumvallate (“surrounded by a wall”) or vallate papillae is arranged in a V-shaped line two thirds of the way back along the dorsal surface of the tongue. Each circumvallate papilla is surrounded by a deep groove in the lingual epithelium (Fig. 13-2A), with about 250 taste buds located in the walls of the groove. Hence, even though the circumvallate papillae are few in number, they contain nearly half of the 5000 taste buds found on an average tongue.
Although an “average” tongue contains about 5000 taste buds, the numbers of both papillae and taste buds are surprisingly variable. For example, among normal, healthy individuals, some have as many as 100 times more taste buds in their fungiform papillae than others do. This is presumably the basis of the 100-fold variation in threshold concentration for various substances among normal individuals.
Taste buds are usually associated with the tongue, but they are also distributed widely, although in smaller numbers, over the palate and pharynx.* The pharyngeal and palatal taste buds are probably more important for swallowing and for reflex responses to good or bad tastes than for conscious awareness of taste.
Taste Receptor Cells Are Modified Epithelial Cells with Neuron-Like Properties
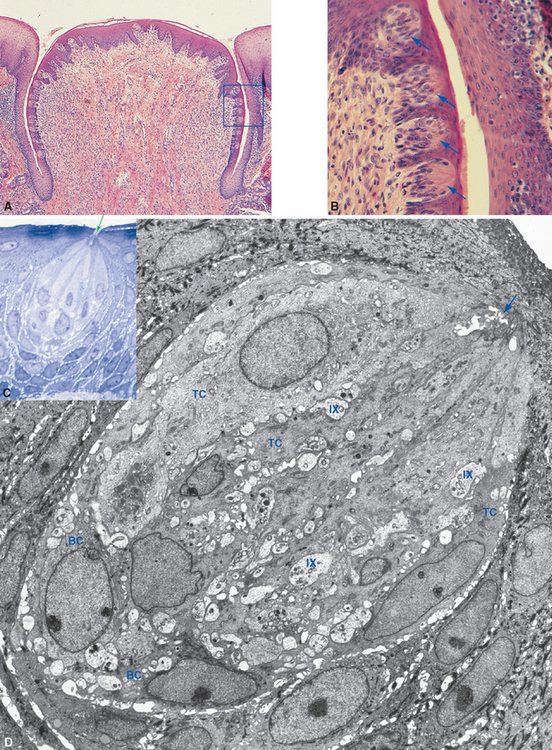
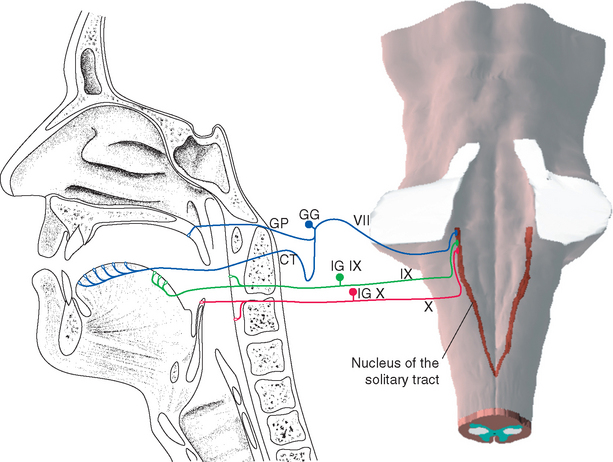
Each taste bud is an ovoid structure containing about 100 spindle-shaped epithelial cells modified as taste cells. Some of these are supporting cells with glial properties, and the rest are taste receptor cells (Fig. 13-2B to D). Taste receptor cells have microvillar processes that extend through a small opening, the taste pore, where they are exposed to chemical stimuli. At the deep end of the taste bud, the receptor cells communicate with visceral sensory fibers from the facial, glossopharyngeal, and vagal nerves (Fig. 13-3). Some make typical chemical synapses, releasing adenosine triphosphate (ATP) and probably other transmitters onto these afferent endings; others use a less conventional communication method, releasing ATP directly into extracellular space, where it diffuses to the same afferent endings. Fibers from the facial nerve innervate the taste buds of fungiform and anterior foliate papillae and the palate; fibers from the glossopharyngeal nerve innervate those of circumvallate and most foliate papillae and the pharynx; and a few vagal fibers innervate those of the epiglottis and esophagus (Fig. 13-4).
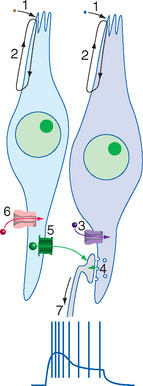
Taste receptor cells differentiate from the surrounding lingual epithelium and subsequently depend on chemical interactions with the gustatory nerves for their continued existence; denervation of an area of tongue causes degeneration of its taste buds. Despite this epithelial origin, taste receptor cells have some very neuron-like properties: they contain transduction machinery in their apical membranes and produce receptor potentials in response to appropriate taste stimuli; some make typical chemical synapses on the peripheral endings of gustatory nerves, and others even produce action potentials when sufficiently depolarized by a receptor potential.* However, unlike almost all neurons, taste receptor cells have a limited life span. Each lives only a week or two before being replaced by differentiation of basal cells, which migrate in from the surrounding epithelium and wait, as their name implies, near the base of the taste bud.
Taste Receptor Cells Utilize a Variety of Transduction Mechanisms to Detect Sweet, Salty, Sour, and Bitter Stimuli
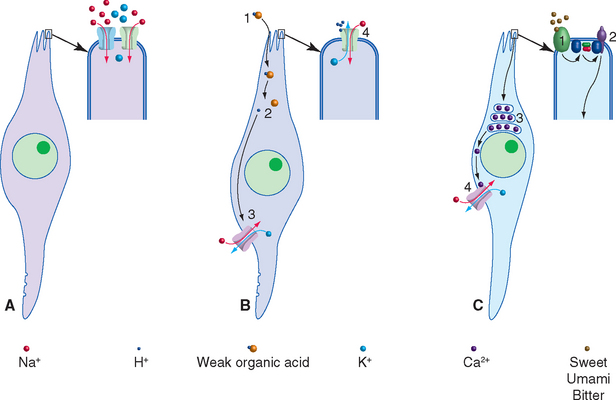
The four basic taste qualities traditionally recognized are sweet, salty, sour, and bitter, but there are others. For example, the flavor-enhancing effect of glutamate (as in monosodium glutamate, or MSG) is a separate taste in its own right. This taste is often referred to as umami, which is Japanese for “delicious,” and it is based on taste receptor cells specifically sensitive to glutamate. Other receptor cells may be selectively sensitive to fats and other components of tastants. Although some areas of the tongue are somewhat more sensitive to certain taste qualities—the tip to sweet, the sides to salt and sour, and posterior portions to bitter—all parts of the tongue are sensitive to all kinds of tastants. Corresponding to these multiple taste qualities, taste receptor cells utilize multiple methods to transduce chemical stimuli into electrical signals (Fig. 13-5). Some are epithelial mechanisms adapted for use in taste transduction, and others are familiar ligand-gated or G protein–coupled mechanisms. Transduction processes include the following:
Second-Order Gustatory Neurons Are Located in the Nucleus of the Solitary Tract
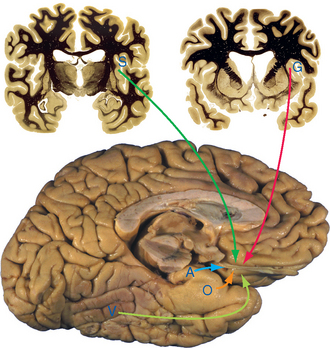
Chemosensory information reaches consciousness as the perception of flavor, but in its role in autonomic responses and the acquisition of food, it has much closer ties to the hypothalamus and limbic system than do other senses. The second-order neurons that mediate the involvement of taste in all these connections are located in the nucleus of the solitary tract (see Fig. 12-11). The nucleus of the solitary tract, the principal visceral sensory nucleus of the brainstem, receives (via the solitary tract) gustatory afferents as well as the other visceral sensory fibers mentioned in Chapters 12 and 23. However, the gustatory fibers and chemosensitive trigeminal fibers end separately in lateral and rostral portions of the solitary nucleus (Fig. 13-4).
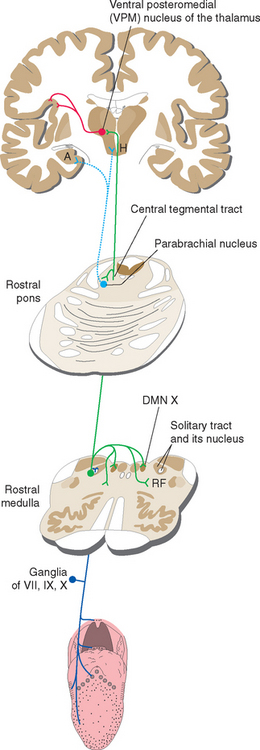
Second-order taste fibers do two things (Fig. 13-6). Some participate in reflex activities, such as salivation, swallowing,* and coughing, by way of cranial nerve motor nuclei. Others, like fibers in most sensory systems, project to the cerebral cortex by way of the thalamus. In this case, however, the projection is uncrossed. Fibers travel through the ipsilateral central tegmental tract to the most medial part of the ventral posteromedial (VPM) nucleus, where they end adjacent to the uncrossed fibers of the dorsal trigeminal tract (see Fig. 12-16). This medial part of VPM then projects to gustatory cortex, which is located in the insula and the medial surface of the frontal operculum, near the base of the central sulcus. Gustatory cortex projects in turn to orbital cortex of the frontal lobe (see Fig. 13-21), where taste information is integrated with olfactory and other information, and to the amygdala, through which taste information reaches the hypothalamus and the limbic system. (In most mammals, taste information reaches the hypothalamus and amygdala more directly, through a projection from the parabrachial nuclei* of the pontine reticular formation, which also distribute nociceptive information and information from visceral structures. The gustatory portion of this set of functions appears to have been lost in primates.)
Information About Taste Is Coded, in Part, by the Pattern of Activity in Populations of Neurons
Sensory systems such as somatic sensation and vision depend on the receipt of information about specific aspects of a stimulus, highly organized in space and time (see Figs. 3-30, 10-20, and 17-35). Activity in many of the neurons in these systems signals the occurrence of a particular kind of event—particularly its location, as is evident in the detailed topographic maps in places such as somatosensory, visual, and motor cortex (see Figs. 3-30 and 17-29). In contrast, the nervous system also uses combinations of inputs to interpret stimuli, as in the case of Ruffini endings that are involved in the sense of touch but produce no sensation when stimulated in isolation (see Chapter 9). Taste and olfaction also utilize this combinatorial type of processing. Taste stimuli are widely dispersed in the mouth during chewing, and odorants fill the nose during breathing, so spatial localization is less important and we actually use the touching of the tongue by a piece of food to localize it (see Box 9-2). Instead, the gustatory and olfactory systems together are faced with the task of coding the identities of many thousands of different chemicals, both singly and in mixtures. In principle, this could be accomplished by having many thousands of different receptor types, each particularly sensitive to one chemical (“labeled-line” coding). An alternative strategy is to code the identity of a stimulus in terms of the pattern of activity in a large population of neurons with multiple sensitivities (“across-fiber” coding). Although this might seem like a cumbersome way to do things, it is actually an efficient system for encoding information about a large variety of potential stimuli using a relatively small number of neurons, similar in principle to using just three kinds of cones as the basis for perceiving hundreds of hues (see Chapter 17).
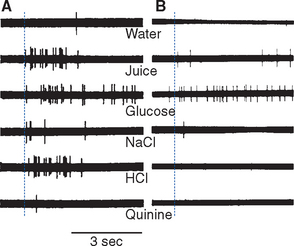
The gustatory system uses a combination of labeled-line and across-fiber coding; the relative importance of the two strategies is still debated. Taste receptor cells apparently contain in their apical membranes only one of the transduction mechanisms shown in Figure 13-5, but at every subsequent level of the gustatory system individual cells respond to more than one kind of tastant. Gustatory nerve fibers branch and innervate multiple taste buds that may even be located in multiple papillae, and ATP released by an assortment of taste receptor cells reaches each afferent ending. As a result, each responds to more than one tastant, although one kind of sensitivity usually predominates for any given nerve fiber. This “tuning” corresponds to the relative sensitivities of different areas of the tongue: most facial nerve (chorda tympani) fibers are maximally sensitive to sweet or salty stimuli, and most glossopharyngeal fibers to sour or bitter stimuli. Second-order neurons in the nucleus of the solitary tract are more broadly tuned (Fig. 13-7A), although there is still some selectivity. By the time gustatory information reaches higher cortical levels, specific features of a taste stimulus have been extracted, and neurons with particular chemical sensitivities are found (Fig. 13-7B).
< div class='tao-gold-member'>

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


