FIGURE 18.1 Section through the medulla at the level of the inferior olivary nucleus.
CN IX exits the skull through the jugular foramen, lateral and anterior to CNs X and XI within a separate dural sheath. After leaving the skull, CN IX enters the carotid sheath, descends between the internal jugular vein and internal carotid artery, dips beneath the styloid process, and then passes between the internal and external carotid arteries. It curves forward, forming an arch on the side of the neck to reach the lateral pharyngeal wall, and then disappears under the hyoglossus muscle to divide into its terminal branches. Two ganglia lie on the nerve just caudal to the jugular foramen: the superior (jugular) and inferior (petrosal) glossopharyngeal ganglia (Figure 18.2). The superior glossopharyngeal ganglion is small, inconstant, has no branches, and is often fused with the inferior ganglion. CN IX has six terminal branches: (a) the tympanic nerve (Jacobson nerve), (b) carotid, (c) pharyngeal, (d) muscular, (e) tonsillar, and (f) lingual branches. CN IX has important connections with CNs V, VII, and X and the cervical sympathetics.
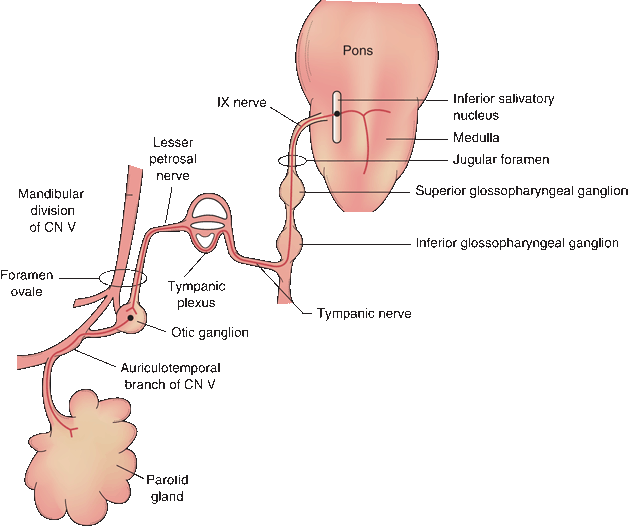
FIGURE 18.2 Peripheral distribution of the parasympathetic branches of the glossopharyngeal nerve.
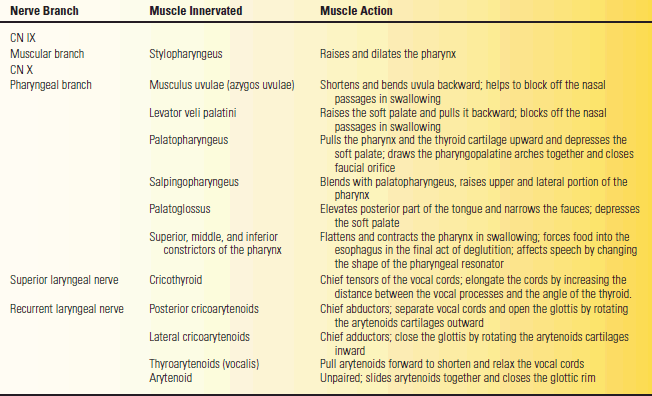
The branchiomotor fibers of CN IX go to the pharynx. The muscular branch follows along the posterior border of the stylopharyngeus muscle, and then terminates in the belly of the muscle. Most of the pharyngeal muscles are supplied by both IX and X. If CN IX supplies any muscle alone, it is the stylopharyngeus. The actions of the stylopharyngeus are described in Table 18.1.
TABLE 18.1 Branches of the Glossopharyngeal and Vagus Nerves, the Muscles Innervated, and Their Actions
CN IX supplies parasympathetic innervation to the parotid gland and to the mucous membranes of the posterior inferior mouth and pharynx (Figure 18.2). The parasympathetic nuclei in the lower brainstem are the superior and inferior salivatory and the dorsal motor nucleus of CN X (DMNX), also known as the dorsal motor or dorsal efferent nucleus of the vagus. The autonomic fibers of CN IX arise primarily from the inferior salivatory nucleus, with some from the DMNX. The parasympathetics pass through the superior and inferior glossopharyngeal ganglia without synapsing. Just below the inferior ganglion, they exit to form the tympanic nerve, which ascends to the tympanic cavity through a small canal on the undersurface of the temporal bone between the carotid canal and the jugular fossa (tympanic canaliculus). The tympanic nerve ramifies in the tympanic cavity to form part of the tympanic plexus. The lesser petrosal nerve is a continuation of the tympanic nerve that leaves the tympanic plexus, enters the middle cranial fossa briefly, and then exits through the foramen ovale to synapse in the otic ganglion. Postganglionic fibers join the auriculotemporal branch of the mandibular division of CN V for distribution to the parotid gland.
Sensory neurons of CN IX are located in the superior and inferior glossopharyngeal ganglia. There are GSA fibers that convey ordinary exteroceptive sensation; GVA fibers that convey information from the carotid body and carotid sinus, as well as visceral sensation from the pharynx; and special visceral afferents that convey taste sensation. The GSA fibers convey exteroceptive sensation from the mucous membranes of the tympanic cavity, mastoid air cells, and auditory canal via the tympanic plexus and tympanic branch. Sensation from the pharynx, tonsil, and posterior third of the tongue travels via the pharyngeal, tonsillar, and lingual branches. Central processes of these cells terminate in the trigeminal nuclei, and their central connections are the same as for other GSA fibers. One of the most important functions of CN IX is to carry visceral afferent fibers from the carotid body and sinus involved in the reflex control of heart rate, blood pressure, and respiration. The carotid branch of CN IX (carotid sinus nerve, sinus nerve of Hering) arises just below the jugular foramen and descends on the internal carotid artery to the carotid sinus and carotid body. It conveys impulses from carotid body chemoreceptors and carotid sinus baroreceptors and terminates centrally on cells in the middle third of the nucleus of the solitary tract. Other fibers carrying visceral afferent fibers from the mucous membranes of the pharynx, soft palate, and posterior third of the tongue pass through the petrous ganglion to terminate in the solitary tract and nucleus. The lingual branches of CN IX carry taste fibers (primarily sour and bitter) from the circumvallate papillae, mucous membranes of the base and taste buds on the posterior third of the tongue, glossoepiglottic and pharyngoepiglottic folds, and lingual surface of the epiglottis. These fibers terminate in the rostral part of the nucleus of the solitary tract (gustatory nucleus). Their central connections are the same as for the taste fibers of CN VII.
Clinical Examination
CN IX is difficult to examine because most or all of its functions are shared by other nerves and because many of the structures it supplies are inaccessible. It is possible to examine pain and touch sensation of the pharynx, tonsillar region and soft palate, and the gag reflex. Bedside testing of taste on the posterior third of the tongue is difficult and seldom attempted. It is not possible to isolate the motor functions from those of the vagus. The small area of cutaneous exteroceptive sensory supply is shared by other nerves. Patients with CN IX lesions might theoretically have detectable sensory loss, but it is not possible to find in patients who have undergone ninth nerve section for glossopharyngeal neuralgia.
The only muscle to receive its motor innervation purely from CN IX is the stylopharyngeus. The only deficit that might be detectable is a slight lowering of the palatal arch at rest on the involved side. Other palatal motor functions are subserved by either CN X or the two nerves working together. The salivary reflex is flow of saliva from the parotid duct after gustatory stimuli. The afferent limb is through taste fibers and the efferent through the parasympathetic outflow of the superior and interior salivatory nuclei.
The gag reflex is elicited by touching the pharynx or palate. Some sources make a distinction between the pharyngeal reflex and the palatal reflex, referring only to the former as the gag reflex. In common clinical usage, no distinction is made between these two and either is referred to as the gag reflex. The reflex is elicited by touching the lateral oropharynx in the region of the anterior faucial pillar with a tongue blade, applicator stick, or similar object (pharyngeal reflex), or by touching one side of the soft palate or uvula (palatal reflex). The pharyngeal reflex is the more active of the two. The reflex also occurs with touching the base of the tongue or posterior pharyngeal wall. The afferent limb of the reflex is mediated by CN IX and the efferent limb through CNs IX and X. The reflex center is in the medulla. The motor response is constriction and elevation of the oropharynx. This causes the midline raphe of the palate and the uvula to elevate and the pharyngeal constrictors to contract. The activity on the two sides is compared. The gag reflex is protective; it is designed to prevent noxious substances or foreign objects from going beyond the oral cavity. There are three motor components: elevation of the soft palate to seal off the nasopharynx, closure of the glottis to protect the airway, and constriction of the pharynx to prevent entry of the substance.
When unilateral pharyngeal weakness is present, the raphe will deviate away from the weak side and toward the normal side. This movement is usually dramatic. For a video of an abnormal gag reflex with palatal deviation, see http://www.youtube.com/watch?v=S9s5ZHCzOXM. Minor movements of the uvula and trivial deviations of the midline raphe are not of clinical significance. In normal adults, both palatal and pharyngeal reflexes are usually present but there may be inter-and intraindividual variation in the intensity of the stimulus required. The gag reflex may be bilaterally absent in some normal individuals. Unilateral absence signifies a lower motor neuron lesion. Like most bulbar muscles, the pharynx receives bilateral supranuclear innervation, and a unilateral cerebral lesion does not cause detectable weakness.
The gag reflex is often used to predict whether or not a patient will be able to swallow. A poor gag reflex in an awake patient with an acute deficit may be a predictor of swallowing difficulties. In fact, the gag reflex has little to do with normal swallowing. Normal deglutition is a smooth coordinated sequence of muscle contractions that propel a bolus of food from the mouth into the esophagus. A normal swallow bears little resemblance to the chaos of a gag reflex. Higher cortical centers have to inhibit the gag response during normal swallowing. The gag reflex is useful but limited in assessing airway protection. A decreased gag reflex in a patient with depressed consciousness may portend inadequate guarding of the airway and increased aspiration risk, but the status of the gag reflex is not a completely reliable indicator. Patients with an apparently intact gag reflex may still aspirate, and a patient with a depressed gag reflex may not.
Davies et al. found the gag reflex absent in 37% of normals, which may explain its low predictive value in the assessment of aspiration risk. Leder and Espinosa concluded that the clinical examination, a major component of which is the status of the gag reflex, underestimated the probability of aspiration in patients who were at risk and overestimated it in patients who were not. The trigeminal nerve contributes to palatal sensation and may allow for paradoxical preservation of the gag reflex in the face of a CN IX lesion.
The gag reflex may be hyperactive in some normal individuals, even to the point of causing retching and vomiting. A hyperactive gag reflex may occur with bilateral cerebral lesions, as in pseudobulbar palsy and amyotrophic lateral sclerosis (ALS).
Disorders of Function
Unilateral supranuclear lesions cause no deficit because of the bilateral corticobulbar innervation. Bilateral supranuclear lesions may cause pseudobulbar palsy (Chapter 21).
Isolated lesions of CN IX are extremely rare if they ever occur. In all instances, the nerve is involved along with other CNs, especially CN X. Nuclear and infranuclear processes that may affect CN IX include intramedullary and extramedullary neoplasms and other mass lesions (e.g., glomus jugulare tumor), trauma (e.g., basilar skull fracture or surgical dissection), motor neuron disease, syringobulbia, retropharyngeal abscess, demyelinating disease, birth injury, and brainstem ischemia. Surgical section or other trauma to the carotid branch may cause transient or sustained hypertension. Involvement of CN IX may be related to the cardiovascular dysautonomia that sometimes accompanies Guillain-Barré syndrome. CN IX may be involved along with other CNs in lesions of the skull base, for example, the jugular foramen syndrome (Chapter 21).
Perhaps the most important lesion of the ninth nerve is glossopharyngeal (or vagoglossopharyngeal) neuralgia or “tic douloureux of the ninth nerve.” In this condition, the patient experiences attacks of severe lancinating pain originating in one side of the throat or tonsillar region and radiating along the course of the eustachian tube to the tympanic membrane, external auditory canal, behind the angle of the jaw, and adjacent portion of the ear. As in trigeminal neuralgia, there may be trigger zones; they are usually in the pharyngeal wall, fauces, tonsillar regions, or base of the tongue. The pain may be brought on by talking, eating, swallowing, or coughing. It can lead to syncope, convulsions, and rarely to cardiac arrest because of stimulation of the carotid sinus reflex. Glossopharyngeal neuralgia must be differentiated from other craniofacial neuralgias and from pain due to a structural lesion of the nerve. Some authorities differentiate between glossopharyngeal neuralgia, in which the pain radiates from the throat to the ear, and Jacobson neuralgia, in which the pain is limited to the ear and eustachian tube. Glossopharyngeal neuralgia is most often idiopathic but has been reported with lesions involving the peripheral distribution of the nerves. Multiple sclerosis only rarely causes glossopharyngeal neuralgia, although it is commonly associated with trigeminal neuralgia.
Carotid sinus hypersensitivity is due to inadvertent activation of the baroreceptors in the carotid sinus causing bradycardia and hypotension. Identifiable etiologies may include constriction around the neck (e.g., tight collar) or a mass in the neck impinging on the sinus, but many cases are idiopathic.
THE VAGUS NERVE
Anatomy and Physiology
The vagus (L. “wandering,” because of its wide distribution) is the longest and most widely distributed CN (Figure 18.3). Some of the nuclei of origin are the same as for CN IX, and it shares many functions with CN IX. It connects with four brainstem nuclei: the nucleus ambiguus, the DMNX, the nucleus of the spinal tract of CN V, and the nucleus of the solitary tract. It conveys exteroceptive GSA sensation from the pharynx, larynx, ear, and meninges and GVA fibers from the larynx, viscera of the thorax and abdomen, and receptors in the aorta. CN X carries skeletomotor axons from the nucleus ambiguus to the pharynx and larynx, and parasympathetic axons from the DMNX to the smooth muscles and glands of the pharynx and larynx and to the thoracic and abdominal viscera. Its terminal ramifications reach the splenic flexure of the colon.
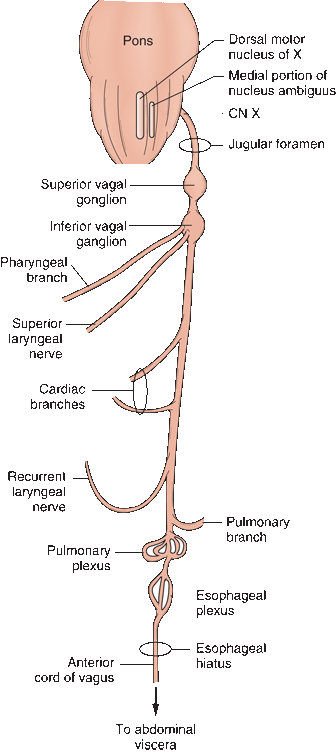
FIGURE 18.3 Peripheral distribution of the branches of the vagus nerve.
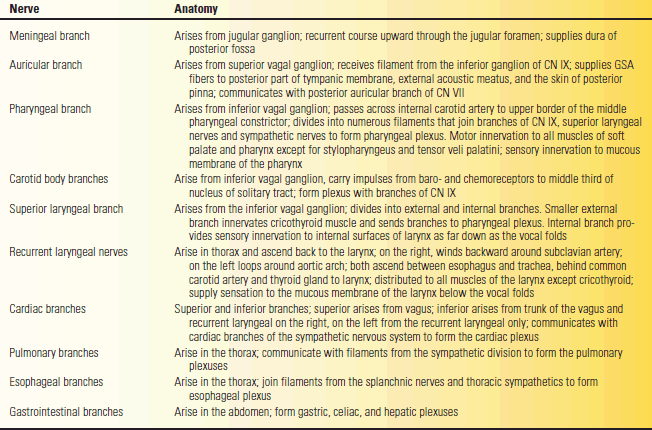
The vagus emerges from the medulla as a series of rootlets just below those of the glossopharyngeal. CN X leaves the skull through the jugular foramen in the same neural sheath as the cranial root of CN XI and behind CN IX. In the jugular foramen, the nerve lies close to the jugular bulb, a dilatation of the internal jugular vein that houses the glomus jugulare (tympanic body). The glomus jugulare has functions similar to the carotid body. CN X descends the neck in the carotid sheath, lying between the carotid artery and internal jugular vein to the upper border of the thyroid cartilage, then between the vein and common carotid to the base of the neck. Branches leave in the jugular foramen to supply the meninges and ear; other branches leave distal to the foramen to supply the pharynx and larynx. The major portion of the nerve enters the thorax. The vagus has two sensory ganglia. The superior (jugular) vagal ganglion is located in the jugular fossa of the temporal bone; the inferior (nodose) ganglion is located just distal to the jugular foramen. There are 10 major terminal branches that arise at different levels: (a) meningeal, (b) auricular, (c) pharyngeal, (d) carotid, (e) superior laryngeal, (f) recurrent laryngeal, (g) cardiac, (h) esophageal, (i) pulmonary, and (j) gastrointestinal. The terminal branches are summarized in Table 18.2.
TABLE 18.2 The Terminal Branches of the Vagus Nerve
The Motor Portion
The cortical center regulating vagus function lies in the lower portion of the precentral gyrus; the supra-nuclear innervation is bilateral but primarily crossed. Fibers descend in the corticobulbar tracts to synapse in the nucleus ambiguus. The vagal branchiomotor fibers follow the same looping intramedullary course as the fibers of CN IX. There are three major branchiomotor branches: pharyngeal, superior laryngeal, and recurrent laryngeal. The actions of the muscles innervated by the vagus are summarized in Table 18.1.
The pharyngeal branch runs between the internal and external carotid arteries and enters the pharynx, where it ramifies to form the pharyngeal plexus. The plexus also receives fibers from the external laryngeal branch, CN IX, and the sympathetic trunk. The vagus, with a contribution from the bulbar portion of CN XI, supplies all the striated muscles of the soft palate, pharynx, and larynx except for the stylopharyngeus (CN IX) and tensor veli palatini (CN V). The muscles and their actions are summarized in Table 18.1.
The superior laryngeal nerve arises distal to the pharyngeal branch and divides into an internal and external branch. The internal branch is primarily sensory. The external branch supplies the cricothyroid. All of the other intrinsic laryngeal muscles are supplied by the recurrent nerves, except for the arytenoid, which may receive some fibers from the internal branch of the superior laryngeal. The recurrent laryngeal nerves both descend deep into the thorax and then loop back to the larynx. On the right, the recurrent laryngeal arches around the subclavian artery; on the left, around the aortic arch. Each nerve gives off cardiac, tracheal, and esophageal branches, ending on each side as the inferior laryngeal nerve to supply the intrinsic muscles of the larynx.
The Parasympathetic Portion
The parasympathetic component of CN X arises from the DMNX, a long cell column just dorsolateral to the hypoglossal nucleus extending from the upper pole of the inferior olive to the lower portion of the medulla. Some parasympathetic neurons lie immediately adjacent in the medial part of the nucleus ambiguus. The neurons in the nucleus ambiguus innervate the heart, and those in DMNX supply the other vagally innervated viscera. The fibers stream ventromedially and merge with the branchiomotor fibers coming from the nucleus ambiguus. The autonomic fibers leave the medulla as preganglionic fibers of the craniosacral division of the autonomic nervous system. They terminate in ganglia close to the viscera they supply and send short postganglionic fibers directly to the muscular and glandular structures they innervate. The vagus is the longest parasympathetic nerve in the body and mediates many important functions, which are discussed in Chapter 45. In brief, a vagal discharge causes bradycardia, hypotension, bronchoconstriction, bronchorrhea, increased peristalsis, increased gastric secretion, and inhibition of adrenal function. The vagal centers in the medulla that control these functions are themselves under the control of higher centers in the cortex and hypothalamus. Inhibition of vagal function produces the opposite effects.
In its course through the thorax, the right vagus nerve gives off pulmonary and esophageal branches, passes through the esophageal opening in the diaphragm posterior to the esophagus, and then divides into gastric and celiac branches. The left vagus also gives off pulmonary and esophageal branches and then enters the abdomen anterior to the esophagus and divides into several gastric branches.
The Sensory Portion
The superior vagal ganglion is located in the upper part of the jugular foramen. It communicates through several delicate branches with the cranial portion of CN XI and with the petrous ganglion of CN IX, with CN VII, and with the superior cervical ganglion. The inferior vagal ganglion lies just beneath the jugular foramen. The cranial root of the CN XI passes through it to join CN X. The inferior ganglion also communicates with CN XII, the superior cervical ganglion, and the loop between C1 and C2. Both vagal ganglia are sensory, containing unipolar neurons that mediate general somatic, special visceral, and general visceral afferents. The branchiomotor and parasympathetic axons pass through the ganglia without synapsing. The superior ganglion primarily conveys somatic sensation, and most of its communication is with the auricular nerve. The inferior ganglion relays general visceral sensation and taste.
The somatic sensory portion of the vagus conveys pain, temperature, and touch sensation from the pharynx, larynx, ear canal, external surface of the tympanic membrane, and meninges of the posterior fossa. In the larynx, GSA fibers from above the vocal folds travel in the internal laryngeal branch of the superior laryngeal nerve; fibers from below the vocal folds travel with the recurrent laryngeal nerve. Visceral afferents follow the same pathways. General sensory fibers from the region of the ear, ear canal, and tympanic membrane travel in the auricular branch (nerve of Arnold). Stimulation of the auricular branch, as by tickling the ear canal, can produce reflex activation of DMNX with coughing, vomiting, and even syncope. The GSA fibers in CN X synapse in the nucleus of the spinal tract of CN V and are relayed to the thalamus and to the sensory cortex.
Fibers carrying GVAs from the pharynx, larynx, vagally innervated viscera, and from baroreceptors and chemoreceptors in the aorta travel over the peripheral processes of neurons in the inferior vagal ganglion. The central processes terminate in the caudal portion of the solitary tract. Collaterals to the reticular formation, DMNX, and other CN nuclei mediate important visceral reflexes and are involved in the regulation of cardiovascular, respiratory, and gastrointestinal function. There are some taste fibers from the region of the epiglottis and arytenoids, which travel with the taste fibers of CN IX to terminate in the rostral solitary tract.
Normal Functions
Normal functions mediated by CNs IX and X include swallowing, phonation, and airway protection and modulation. The complex process of swallowing is divided into two stages, controlled primarily by CNs IX, X, and XII. In the first stage, the food bolus is driven back into the fauces by tongue action. During the second stage, the epiglottis closes over the entrance to the larynx, and the bolus glides along its posterior surface. The muscles of the soft palate and nasopharynx close above the bolus to prevent passage into the nasopharynx. The bolus is directed downward and backward into the pharynx, and then the constrictors contract to propel it downward into the esophagus.
The larynx is composed of several cartilages. The thyroid and cricoid cartilages form part of the outer casing. The arytenoids are paired cartilages lying in the interior; they have a muscular process and a vocal process. The true vocal cords are mucous membranes that cover the vocal ligaments, which extend from the vocal processes of the arytenoids to the thyroid cartilage. The larynx is controlled by myriad small muscles. The arytenoids may either slide or pivot; either action changes the configuration of the vocal cords. The glottic rim is the passageway between the vocal cords. Contraction and relaxation of the intrinsic laryngeal muscles change the tension or shape of the vocal cords and alter the aperture of the glottic rim. The muscles of the larynx perform three basic functions: They abduct and open the glottic rim to allow air entry and exit, they adduct and close the glottic rim to protect the airway during swallowing, and they regulate the tension on the vocal cords to allow phonation. The cricothyroids, posterior and lateral cricoarytenoids, and thyroarytenoids are paired muscles. The arytenoid is unpaired. The actions of the intrinsic laryngeal muscles are summarized in Table 18.1 and Figure 18.4.
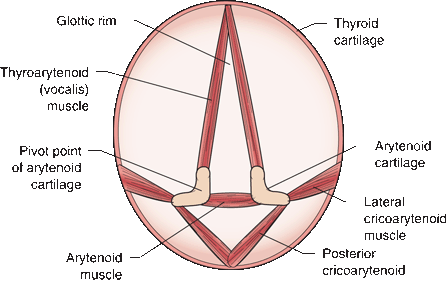
FIGURE 18.4 The cricothyroid muscles (not shown) tilt the thyroid cartilage forward on the cricoid cartilage, tensing the vocal cords. The thyroarytenoid muscles run from the thyroid cartilage to the arytenoid cartilages; contraction tenses the vocal cords. The other muscles attach to the cricoid cartilage. The paired arytenoids may either slide or pivot. Contraction of the arytenoid muscle pulls the arytenoid cartilages together, adducting the cords and closing the glottic rim. The lateral cricoarytenoid muscle causes the vocal process of the arytenoid to pivot medially, adducting the cords. The posterior cricoarytenoid causes the vocal process to rotate laterally, abducting the cords.
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


