FIGURE 24.1 The relationship of the spinal cord segments and spinal nerves to the vertebral bodies and spinous processes.
In newborns, the spinal cord may extend as far caudally as L3. During maturation, the vertebral column elongates more than the spinal cord, and the adult spinal cord is about 25 cm shorter than the vertebral column. The variability of the lower level has some correlation with length of trunk, especially in females. Radiographically, the conus is usually seen at the L1-L2 interspace in adults; if the level of the tip of the conus is below the mid-L2 vertebral body, the conus is considered low-lying. Since the spinal cord normally ends at the level of the L1-L2 interspace, lumbar punctures are done well below this level.
The length discrepancy between the spine and the spinal cord creates a difference between the segments of the spinal cord and the vertebral level that progressively increases from rostral to caudal. In the upper cervical area, the numerical cord level is about one segment greater than the corresponding vertebral spinous process (e.g., the C5 spinous process lies at the C6 segment of the spinal cord). In the lower cervical and thoracic areas, there is a difference of about two segments; in the lumbar region, there is a difference of almost three segments (Figure 24.1). Because of the offset, the spinal nerves below the cervical region course downward before exiting through the intervertebral foramina. Nerve roots exit through the foramen above the vertebra of like number from C1 to C7. The C8 root exits below C7 and sets the pattern of root exit below like-numbered vertebra followed down the rest of the vertebral column. The lumbar and sacral roots descend almost vertically to reach their points of exit. These long trailing roots from the lower cord segments make up the cauda equina.
The spinal cord is greater in width and diameter in the cervical and lumbosacral regions, forming the cervical and lumbar enlargements, the site of nuclear centers that supply the extremities. The cervical enlargement extends from the C3–T2 spinal cord segments; it innervates muscles of the upper limb (Figure 24.2). The lumbar enlargement extends from the L1–S3 spinal cord segments; it innervates muscles of the lower limb. The segments of the cervical enlargement match fairly well with the corresponding vertebral levels. The lumbar enlargement extends over vertebral levels T9-T12. Below T12, the spinal cord tapers to form the conus medullaris.
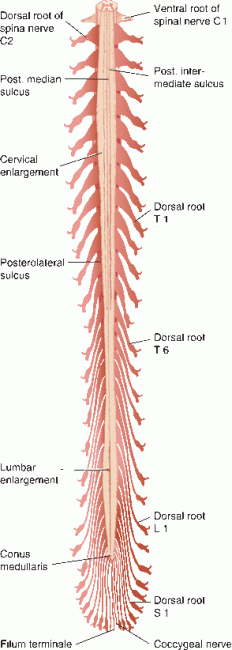
FIGURE 24.2 Posterior view of spinal cord showing attached dorsal root filaments and spinal ganglia. Letters and numbers indicate corresponding spinal nerves. (Modified from Carpenter, MB, Sutin, J. Human Neuroanatomy. Baltimore, MD: Williams & Wilkins, 1983; courtesy of Lippincott Williams & Wilkins.)
Each segment of the spinal cord gives rise to a mixed spinal nerve that contains motor, sensory, and autonomic fibers (Figure 24.3). Motor axons arising from the anterior horn cells of the spinal cord travel in the converging filaments of the anterior spinal root. On each posterior root, within the intervertebral foramen and just proximal to the junction with the anterior root, lies a DRG. The DRG is made up of unipolar neurons, and the posterior roots are made up of the central processes of these neurons. Acetylcholine is the only neurotransmitter in the anterior roots; the posterior roots contain several, including substance P, glutamate, calcitonin gene-related peptide, vasoactive intestinal polypeptide, cholecystokinin, somatostatin, and dynorphin. The anterior roots convey motor and autonomic fibers into the peripheral nerve; they join the posterior root to form the mixed spinal nerve. In the thoracolumbar region, white and gray rami communicantes connect the spinal nerve to the paravertebral sympathetic chain (Figure 24.3).
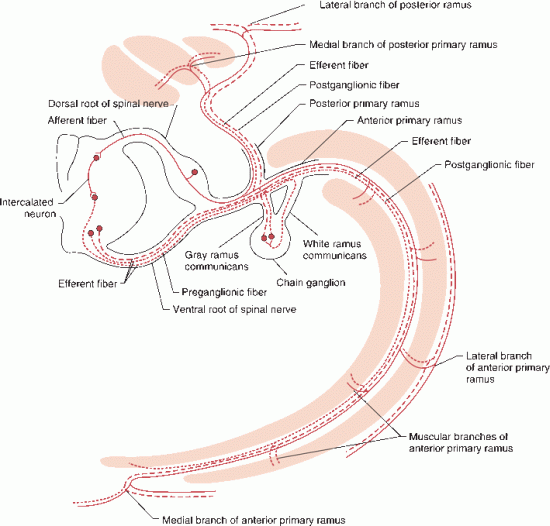
FIGURE 24.3 Segmental spinal nerve showing the course of motor, sensory, and preganglionic and postganglionic sympathetic fibers.
The Bell-Magendie law (for Sir Charles Bell and Francois Magendie) states that the anterior roots are motor and the posterior roots sensory; this discovery was one of the seminal developments in early neurobiology. However, it now appears there may be some afferent nerve fibers in the anterior roots, and up to 3% of the fibers in the posterior root may be efferent. The roots pass through the dura separately and then unite in the intervertebral foramina just distal to the DRG to form the mixed spinal nerve. After the mixed spinal nerve exits the intervertebral foramen, it divides into anterior and posterior primary rami. The smaller posterior primary rami supply the skin of the back and the paraspinal muscles. The anterior primary rami are continuations of the mixed spinal nerves and supply motor and sensory innervation to all other structures of the body. The anterior primary rami from the cervical and lumbar enlargements form the brachial and lumbosacral plexuses that innervate the limbs. The anterior primary rami of the thoracic segments of the spinal cord continue as the intercostal nerves. The anterior primary ramus of the mixed spinal nerve is sometimes referred to as a root, especially by surgeons and especially regarding the brachial plexus. When anatomy sources say that the C5 and C6 roots join to form the upper trunk, they are actually referring to the anterior primary rami of the spinal nerve. Similarly, when the dorsal scapular nerve is said to arise from the C5 root.
The delicate pia-arachnoid closely invests the spinal cord. The tough, fibrous membrane of the dura mater forms a firm, tubular sheath around the exiting nerve roots. The spinal cord is separated from the walls of the vertebral canal by the epidural space, which contains areolar tissue and a plexus of veins. The subdural space is a potential space containing a small amount of fluid. The subarachnoid space is a well-defined cavity containing cerebrospinal fluid that extends to about the level of the second sacral vertebra, forming the lumbar thecal sac. Systemic malignancies frequently metastasize to the capacious spinal epidural space. Spinal hematomas may accumulate in the epidural, subdural, or subarachnoid space. Subarachnoid hematomas can extend along the entire length of the subarachnoid space. Epidural and subdural spinal hematoma present with intense, knife-like pain at the location of the hemorrhage (“coup de poignard”) followed by progressive paralysis below the affected level.
A section of spinal cord where the rootlets of a pair of spinal nerves attach is referred to as a segment, although there is no internal demarcation to separate one segment from another. Each spinal cord segment can function as an independent entity for purposes of some very basic functions, such as the segmental muscle stretch reflex. Each segment controls the resting muscle tone of the muscles it innervates. The motor units supplying the myotomal muscles innervated by a segment carry out voluntary activity. The motor function of a spinal cord segment is modulated and influenced by suprasegmental impulses from several descending motor tracts.
The spinal cord parenchyma consists of an H-or butterfly-shaped core of gray matter that contains nerve cells, surrounded by white matter made up of longitudinally arranged ascending and descending nerve fibers, mainly myelinated. The relative proportion of white to gray matter varies depending on the cord level. In the center of the gray matter, running throughout the entire length of the cord and for a short distance into the filum terminale, is a minute central canal consisting of a single layer of ependymal cells. The two halves of the spinal cord are joined by a commissure made up of a core of gray matter surrounded by anterior and posterior white commissures. Internally, the white matter of the spinal cord is divided into posterior, lateral, and anterior funiculi. The posterior funiculus extends from the posterior median sulcus to the attachment of the posterior rootlets at the posterolateral sulcus. Rostral to the upper thoracic segments a posterior intermediate sulcus separates the medial fasciculus gracilis from the lateral fasciculus cuneatus. The lateral funiculus lies between the attachments of the posterior and anterior spinal rootlets. The anterior funiculus extends from the anterior rootlets to the anterior median fissure.
The spinal cord gray matter consists of anterior and posterior horns with a lateral concavity. In the thoracic and upper lumbar regions, an intermediolateral column of autonomic neurons forms a small projecting lateral horn between the anterior and posterior horns. The sympathetic axons project through the anterior horn and anterior root and then through the gray rami communicantes to enter the sympathetic chain ganglia. The gray matter contains neurons, nerve fibers, supporting neuroglia, and blood vessels. Neurons are not distributed uniformly but are collected into functional groups that consist of columns of cells extending over many segments (Figure 24.4). The most elementary division is into posterior horns that contain sensory neurons and anterior horns that contain motor neurons. The posterior horn is relatively narrow and capped by a thin crescent of tissue, the substantia gelatinosa (of Rolando). The tip of the posterior horn is separated from the surface by a thin white matter tract, the dorsolateral tract (of Lissauer).
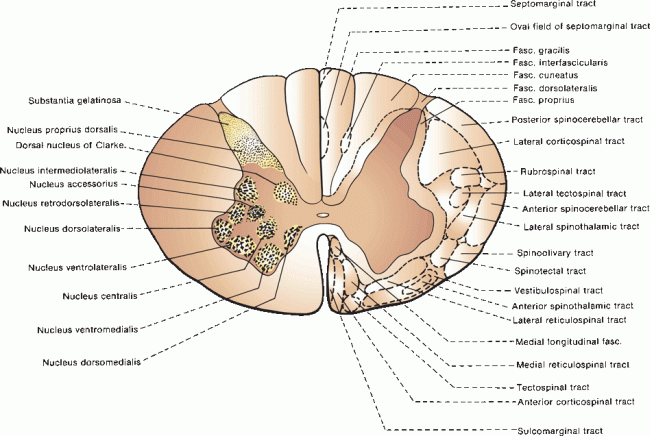
FIGURE 24.4 Cross section of the spinal cord showing the arrangement of cellular groups in the gray matter and fiber pathways in the white matter.
Within the anterior horn, there are alpha motor (skeletomotor) neurons, gamma motor (fusimotor) neurons, beta motor neurons, and interneurons. The alpha motoneurons innervate common, extrafusal striated skeletal muscle; the gamma motoneurons innervate intrafusal, muscle spindle fibers (L. fusus “spindle”). Beta motor neurons innervate both intrafusal and extrafusal fibers. All of these fibers are classified as general somatic efferent. At any given level, there is a somatotopic arrangement of motor neurons. A medial cell group that extends throughout the length of the spinal cord innervates trunk and proximal muscles. A lateral cell group found only in the cervical and lumbar enlargements innervates limb muscles. The expansion of the anterior horns in the cervical and lumbar enlargements reflects the presence of this lateral cell column that supplies limb muscles. In both the cervical and lumbar enlargements, neurons innervating proximal muscles are located more rostrally and those innervating distal muscles more caudally; cells supplying extensors are more ventral than cells innervating flexors. Some motor neurons are aggregated into well-defined nuclear groups. The phrenic nucleus is a central collection of cells from C3 to C7 that innervates the diaphragm. Onuf’s nucleus is a ventrolateral cell group at S1 and S2 that supplies the striated muscles of the perineum. For unknown reasons, the cells of Onuf’s nucleus are relatively spared in motor neuron disease but disproportionately involved in multiple system atrophy.
The collections of neurons in the gray matter of the spinal cord are not as well defined as nuclei in other parts of the nervous system. In experimental animals, Rexed identified 10 regions, or laminae, of the spinal cord gray matter. Rexed laminae are more commonly used in the study of spinal cord neurons than the named nuclei (Figure 24.5). Lamina I covers the most posterior part of the posterior horn, lamina IX lies at the most anterior part of the anterior horn, and lamina X surrounds the central canal. There is evidence supporting the laminar scheme in humans as well. Rexed lamina IX contains the motor neurons that supply striated muscles. Small neurons tend to innervate type S (slow twitch, fatigue resistant) motor units, and larger neurons type FF (fast twitch, fatigue sensitive) and FR (intermediate, fast twitch but more fatigue resistant then FF) motor units. Laminae I to IV make up most of the posterior horn and receive cutaneous primary afferents. Laminae V and VI, at the base of the posterior horn, primarily receive muscle spindle afferents as well as fibers from descending suprasegmental motor tracts. Lamina VII includes the intermediolateral gray column and the nucleus dorsalis (of Clarke). Lamina VIII has extensive connections with adjoining laminae and with the opposite side of the spinal cord. The Rexed laminae are described in more detail in Box 24.1; Figure 24.1.
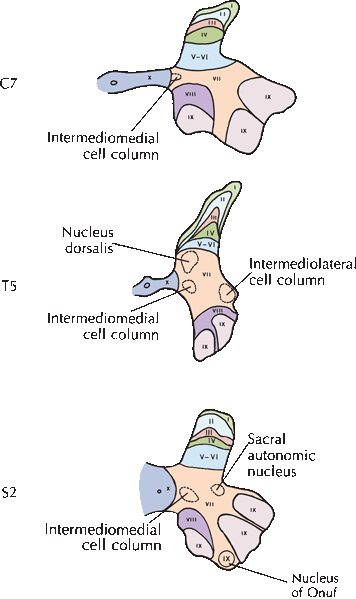
FIGURE 24.5 Positions of the Rexed cytoarchitectonic laminae at three levels of the spinal cord gray matter. (Modified from Kiernan JA. Barr’s The Human Nervous System: An Anatomical Viewpoint. 9th ed. Philadelphia: Wolers Kluwer/Lippincott, Williams & Wilkins, 2009; with permission.)
The Rexed Laminae
Lamina I consists of scattered large neurons in the superficial region of the posterior horn that receive afferents from Lissauer’s tract; these neurons are primarily involved with pain impulses. Lamina II, with overlap into lamina III, is approximately coterminous with the substantia gelatinosa. These regions contain interneurons that receive pain and temperature afferents from Lissauer’s tract but make no contribution to the long ascending sensory pathways. Lamina IV contains polymodal, or wide dynamic range, sensory neurons that are activated by many different stimuli. Cells in lamina IV give rise to the contralateral spinothalamic tract. Laminae V and VI are indistinguishable in humans. Cells in laminae IV and V to VI make up the nucleus proprius. Laminae V to VI receive many descending fibers, particularly those of the corticospinal tract. Lamina VII is the most extensive cytoarchitectonic layer. Between C8 and L3, the medial portion of lamina VII contains the nucleus dorsalis (of Clarke), which is the origin of the posterior spinocerebellar tract. Laterally in lamina VII is the intermediolateral gray column that contains preganglionic sympathetic neurons between T1 and L2 and preganglionic parasympathetic neurons between S2 and S4. Lamina VIII contains interneurons involved in motor control, including Renshaw cells that receive collaterals from nearby lamina IX motor neurons. Lamina IX contains the motor neurons. There is a medial nuclear group at all levels that innervates axial and proximal muscles, and a lateral nuclear group in the cervical and lumbar enlargements that innervates distal extremity muscles. Lamina X surrounds the central canal and contains small neurons that receive dorsal root afferents involved with pain, temperature, and visceral sensation.
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


