10 The thalamus and hypothalamus
 Clinical cases for thought
Clinical cases for thoughtAnatomy of the thalamus
The diencephalon encloses the third ventricle and includes the thalamus with its lateral and medial geniculate bodies, the subthalamus, the epithalamus, and the hypothalamus. Each cerebral hemisphere contains a thalamus, which is a large egg-shaped mass of grey matter, in the dorsal portion of the diencephalon (Fig. 10.1). The thalamus is an important link between sensory receptors and cerebral cortex for all modalities except olfaction.
 Quick facts 10.1
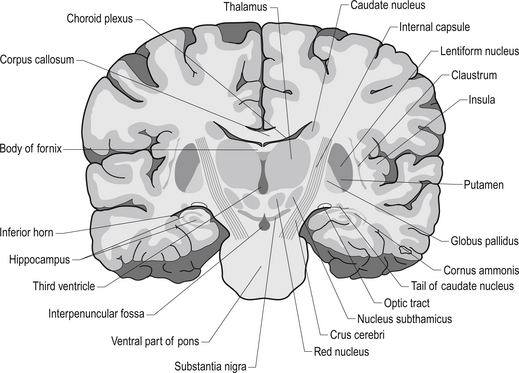
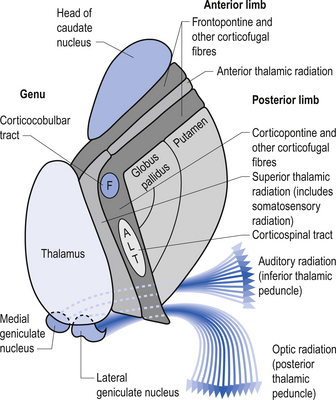
Quick facts 10.1The rostral end of the thalamus, also known as the anterior tubercle, is narrower than the posterior portion of the thalamus, which contains a medial enlargement referred to as the pulvinar and a lateral enlargement referred to as the lateral geniculate body. The medial surface of the thalamus forms the lateral wall of the third ventricle and forms a connection to the medial surface of the opposite thalamus through a short communicating projection of grey matter called the massa intermedia or the central thalamic adhesion (Chusid 1982). The thalamus receives extensive projections from all of the main subcortical areas of the nervous system including spinal cord, hypothalamus, cerebellum, and the basal ganglia and forms reciprocal projections with the majority of the cerebral cortex. The connections to and from the cortex, also known as the thalamic radiations, are carried in four fibre tracts referred to as peduncles or stalks. These projections form a considerable portion of the internal capsule (Fig. 10.2). The anterior thalamic peduncle carries projection fibres from the anterior and medial thalamic nuclei to all areas of the frontal cortex. The superior peduncle carries projection fibres to and from the ventral and lateral thalamic nuclei to the pre- and postcentral gyri and premotor and presensory areas of the cortex. The posterior peduncle caries projection fibres to and from the posterior and lateral thalamic areas including the lateral geniculate body and the pulvinar to the posterior and occipital cortical areas. The inferior thalamic peduncle connects the posterior thalamic areas including the medial geniculate body to the temporal areas of cortex (Williams & Warwick 1984). The external medullary laminae are layers of myelinated fibres on the lateral surface of the thalamus immediately adjacent to the internal capsule (Fig. 10.3). The internal medullary lamina is a vertical sheet of white matter deep in the substance of the thalamus that bifurcates in the anterior portion of the thalamus to divide the substance of the thalamus into lateral, medial, and anterior segments (Fig. 10.3). The thalamus has seven groups of nuclei organised with respect to the internal medullary lamina. These include the anterior nuclear group located rostrally, the nuclei of the midline, the medial nuclei, the ventral nuclei, lateral nuclear mass which expands posteriorly to include the pulvinar, the intralaminar nuclear group, and the reticular nuclei (Fig. 10.3). Several nuclei of the thalamus are considered to be areas of singularity dependent on neural activation from the cortex to survive. These nuclei show marked transneural degeneration if the areas of cortex that project to them are damaged, understimulated, or subject to excessive inhibition (Williams & Warwick 1984). This process is an example of diaschisis, where reduced output from one area of the neuraxis results in degeneration of the downstream neuron pools.
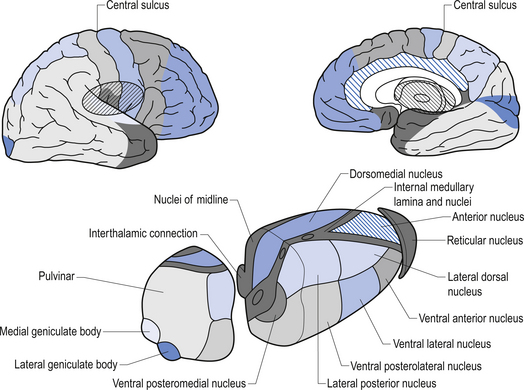
The anterior nuclear group receives input from the ipsilateral mammillary nuclei of the hypothalamus via the mammillothalamic tract and from the presubiculum of the hippocampal formation (Fig. 10.4). Neurons in the anterior thalamic group project to regions of the cingulate and frontal cortices, mainly areas 23, 24, and 32. The anterior group of nuclei is a principal limbic component in linking the hippocampus and the hypothalamus and is involved with the modulation of memory and emotion.
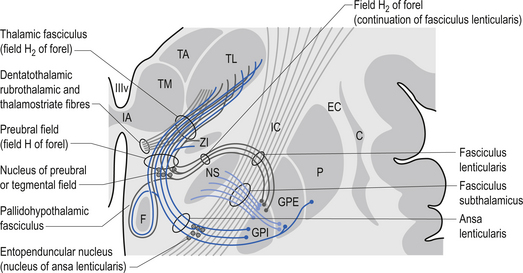
The ventral nuclear group is composed of three nuclei, the ventral anterior (VA), the ventral posterior (VP), and the ventral intermedius (VI). The ventral posterior nucleus is further divided into the functionally important ventral posterior lateral (VPL) and ventral posterior medial (VPM) nuclei. The vast majority of the fibres reaching the ventral group are from the afferent fibres of the sensory system of humans. The VPL receives projections from the contralateral cuneate and gracile nuclei via the medial lemniscal pathway and both contralateral and ipsilateral spinothalamic projections via the anterolateral system. The VPM receives projections from the trigeminal and gustatory lemnisci. These nuclei project reciprocally via the posterior limb of the internal capsule to the somatosensory areas including areas 1, 2, and 3 of cortex (see Fig. 10.2). The VI nuclei receive extensive projections from the dentate and interpositus nuclei of the cerebellum and from the basal ganglia. The VI projects to other thalamic nuclei and to the motor areas of cortex, namely areas 4 and 6. The VA nucleus receives extensive projections from the globus pallidus via the thalamic fasciculus and from the dentate nucleus of the cerebellum. The VA nucleus is therefore very important in the integration or modulation of projections from the basal ganglia and the cerebellum on the cortical areas of motor function.
The reticular nuclei form an outer shell around the lateral aspects of the thalamus. All afferent and efferent projection fibres, to and from the thalamus, pass through this reticular nuclear area. The neurons of this nucleus are predominantly GABA-ergic in nature, while other thalamic nuclei are mainly excitatory and glutaminergic. The reticular nuclei appear not to have direct projections to the cortex but only to other nuclei of the thalamus (Destexhe & Sejnowski 2003).
The lateral geniculate nucleus (LGN) appears as a swelling on the rostral surface of the pulvinar and receives afferent input from the axons of the retinal ganglion cells of the temporal half of the ipsilateral eye and the nasal half of the contralateral eye. The LGN neurons then project axons to the ipsilateral primary visual cortex via the optic radiations. The nucleus consists of six layers of nerve cells and is the terminus of about 90% of the fibres of the optic tract. The remaining 10% of fibres terminate in the pretectal areas of the mesencephalon, the superior colliculus of the tectum of the mesencephalon, and some fibres synapse directly on neurons in the hypothalamus (Snell 2001). Only 10–20% of the projections arriving in the LGN are derived directly from the retina. The remaining projections arise from the brainstem reticular formation, the pulvinar, and reciprocal projections from the striate cortex. These projections between the LGN and the striate cortex are important for a number of reasons but may play a major role in the process of physiological completion or ‘fill-in’ that occurs during visual processing in the cortex.
The medial geniculate nucleus (MGN) or body is the tonotopically organised auditory input to the superior temporal gyrus. It appears as a swelling on the posterior surface of the pulvinar. Afferent fibres arriving in the medial geniculate body from the inferior colliculus form the inferior brachium. The inferior colliculus receives projections from the lateral lemniscus. The MGN receives auditory information from both ears but predominantly from the contralateral ear. The efferent projection fibres of the MGN form the auditory radiations that terminate in the auditory cortex of the superior temporal gyrus (Snell 2001).




