Chapter 49 Ultradian, Circadian, and Sleep-Dependent Features of Dreaming
Abstract
Introduction
Previous reviews of dreaming and chronobiology1,2 concluded that strikingly little convergence had occurred between chronobiology and the study of dreaming following publication of Aserinsky and Kleitman’s work3—despite the substantial accumulations of research in both domains. This chapter focuses on new findings and formulations describing potential ultradian, circadian, and sleep-dependent influences on dreaming and evidence for their interactions.4,5 The term dreaming is used in an inclusive sense equivalent to that of sleep mentation, that is, the occurrence of any subjectively experienced cognitive events during sleep.
Ultradian Factors
Frequency and Length of Recalled Dreams
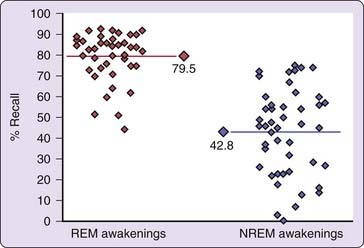
Just as the regular alternation between rapid eye movement (REM) and non-REM (NREM) sleep is thought to be governed by a 90-minute ultradian oscillator, so to do numerous studies support the notion that the amount and intensity of dream mentation fluctuates between a high in REM sleep and a low in NREM sleep. Figure 49-1 shows that peak dream recall (~80%) occurs from REM sleep whereas the lowest level of recall is from NREM sleep (~43%). Paralleling these differences are similarly large REM–NREM differences in dream report length; REM to NREM ratios in total recalled content (TRC) vary from 2 : 1 to 5 : 1.6
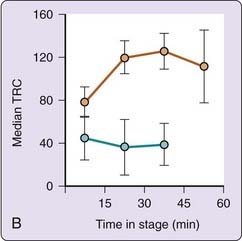
Beyond such dichotomous REM–NREM differences, the oscillatory nature of dream production becomes evident when it is sampled at multiple points within REM or NREM sleep stages. Figure 49-2 (left panel) shows that the length of dream reports, as reflected in TRC, fluctuates sinusoidally as a function of time spent in NREM sleep (blue bars) and time spent in REM sleep (red bars). For NREM sleep, reports are longest from 0 to 15 minutes and from 45 to 60 minutes into stage and shortest in between; for REM sleep the opposite pattern exists.7 A similar sinusoidal fluctuation was found in a replication study (see Fig. 49-2, right panel).6
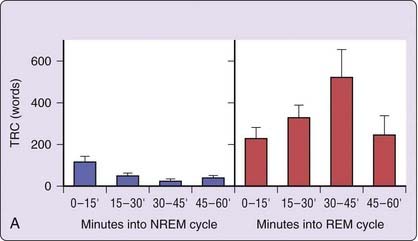
Figure 49-2 Dream report length following awakenings from NREM and REM sleep periods of different durations in two studies. A, Mean (±SEM) report length as a function of elapsed time in stage for 88 REM and 61 NREM reports. TRC, total recall count.
(From Hobson JA, Pace-Schott E, Stickgold R. Dreaming and the brain: towards a cognitive neuroscience of conscious states. Behav Brain Sci 2000;23:793-842.);
Other research supports these findings. Four separate studies8–11 showed NREM dream reports to be either more prevalent or longer when awakenings took place close to a prior REM sleep episode (5 minutes) rather than far from one (10 minutes, 30 minutes, 12 minutes, and 15 minutes, respectively). A fifth study12 showed that NREM sleep duration preceding an awakening was negatively correlated with report length.
Rosenlicht13 reported that report lengths following awakenings from REM periods of 5-minute durations were marginally shorter (P = .114) than those of 10-minute durations. However, given the close proximity of these samples (5-minute difference) on the 90-minute ultradian cycle, such a trend remains consistent with the proposed ultradian oscillator.
In general, dream recall and report length findings support the possibility that dream imagery is determined by the natural variation of an imagery generator oscillating through REM and NREM sleep on a 90-minute frequency. If so, stricter dream sampling criteria that more consistently control for phase relationships between REM and NREM sampling points are needed to clearly demonstrate the relationship. On one hand, awakenings following a consistent delay for both stages (e.g., 10 minutes into the stage) may bias the size of differences between the two states. For example, using the results plotted in Figure 49-2 (left panel), awakenings conducted at 0 to 15 minutes post–stage-onset would clearly lead to a modest 2 : 1 ratio in REM : NREM word count (~200 words vs. 100 words), whereas awakenings conducted at 30 to 45 minutes post–stage-onset would lead to an enormous 20 : 1 difference (~500 words vs. 25 words). On the other hand, the common method of conducting each of several awakenings of the night progressively later into the target stage (e.g., 5 minutes into REM 1, 10 minutes into REM 2, 15 minutes into REM 3, etc.), confounds ultradian phase with circadian and sleep-dependent fluctuations (see later). More accurate assessments of the ultradian dreaming process will require experimental designs sensitive to these confounds as well as the implementation of protocols capable of separating ultradian, circadian and sleep-dependent factors, for example, forced desynchrony and ultrashort sleep–wake protocols (see later).
Quality of Dream Reports
Evidence also indicates that dream qualities such as vividness, intensity, and dreamlikeness oscillate with an ultradian frequency within and between REM and NREM sleep. Many studies (see reviews in Nielsen14 and Hobson et al.15) demonstrate that REM sleep reports are more perceptual, hallucinatory, emotional, dramatic, physically involving, and rich with characters and visual scenes than are NREM reports, whereas the latter are more conceptual, thoughtlike, and mundane.16
However, because REM reports are also consistently longer than are NREM reports, some argue that comparing the two is valid only if this difference is statistically controlled by, for example, selecting equal-length reports, calculating proportions with a common metric, or removing report length as a co-variate. Such procedures have been criticized on methodological grounds,14,15,17 but there is nonetheless consistent evidence that qualitative REM-NREM differences are maintained even after report length is controlled (see review in Nielsen14). Even with length controls, REM dream reports surpass NREM dream reports on measures of emotional intensity,18 self-reflectiveness,19 bizarreness,20,21 visual and verbal imagery,20,22,23 movement imagery,24 characters and self-involvement,25,26 self-representation,26 psycholinguistic structure,27 and narrative linkage.28
Clear within-stage sinusoidal variations of such qualitative measures are more difficult to demonstrate. Dream reports from “long” REM sleep episodes (9 minutes or more) are, relative to those from “short” episodes (1 minute or less), more active, distorted, dramatic, emotional, anxious, unpleasant, and vivid and contain more different scenes, more scenes with clear visualization, and more violence and hostility.16 Similar results were obtained from a small sample (N = 4) of male students each awakened 12 times—twice each from REM 2 and REM 4 for each of six REM onset time delays: 0.5, 2.5, 5.0, 10, 20, and 30 minutes. Of 12 qualities rated, emotion, anxiety, pleasantness, and clarity all showed linear increases over time; emotion, anxiety, and pleasantness showed additional trends suggesting ultradian modulation with peaks at 10 and 30 minutes.29–31
For NREM sleep, two studies suggest an ultradian oscillation opposite to that in REM sleep. In one study,32 dreamlike fantasy scale scores were lower (P < .10) for reports from 20-minute NREM (stage 4) episodes than they were for 5-minute NREM episodes matched within subjects and for time of night. In a second study,10 NREM (stage 2) reports obtained from 12-minute episodes after the end of REM sleep episodes were rated as less dreamlike than were the NREM reports obtained 5 minutes after REM sleep episodes (P < .001).
Additional studies suggest that the types of memory associations that subjects produce as likely sources of their dreams oscillate with ultradian frequency. These are primarily biographical episodes (episodic memories) for NREM dream content, and a mixture of episodic and semantic memories for REM dream content.33–35 The predominance of episodic sources for NREM dreams is independent of time of night and of corrections for report length.33,35–38
In contrast to much of the preceding, variables such as plausibility and sensibleness of the dream do not vary with time-in-stage.39,40 A more exact determination of which qualitative dream features oscillate and which do not may provide clues as to the functional dynamics of dream imagery.
Oscillatory transitions between and within REM and NREM dreaming are both clearly paralleled by regular physiological oscillations, e.g., by ultradian-determined variations in REM sleep propensity. Within-stage changes include variations in EEG power, and autonomic and hormonal measures. The clearest variations occur in stage 2 sleep: autonomic activity increases for stage 2 sleep that precedes REM sleep and decreases for stage 2 sleep that precedes SWS.41 Similarly, fast EEG events such as arousals and stage 2 cyclic alternating pattern A2 and A3 phases often begin well before REM sleep.42 Even more basic regulatory systems, like pontine REM-on neurons, demonstrate a graduated oscillation that begins well before EEG-defined REM sleep onset.43 Such variations led to the speculation41 that stage 2 sleep is fundamental to the ultradian oscillatory process of sleep deepening and lightening. I have linked such changes to the gradual and imperceptible onset and offset of REM sleep processes (“covert REM processes”12,14), but they may equally well be considered ultradian variations in REM sleep propensity that reflect ultradian oscillations in the presence and intensity of dream imagery.
Circadian and Sleep-Dependent Factors
Purely circadian features of dream production are difficult to ascertain because their measurement is usually limited to the nocturnal portion of the sleep–wake cycle and because across-the-night changes that are identified could be due to sleep-dependent processes, circadian influences, or a combination of the two. How might sleep-dependent and circadian influences on dreaming be distinguished? As suggested in the previous section, a useful heuristic is to use the close ultradian coupling of dreaming and REM propensity to evaluate across-the-night changes in dreaming. Using a forced desynchrony protocol, sleep-dependent and circadian-driven patterns of REM sleep propensity (REM%) have been isolated (Fig. 49-3).44 Figure 49-3 (panel A) shows that circadian-driven fluctuations in REM% are characterized by abrupt “switch-like”45 transitions, that is, rapid increases in the middle of the night, whereas sleep-dependent changes (panel B) are gradual and linear in nature.
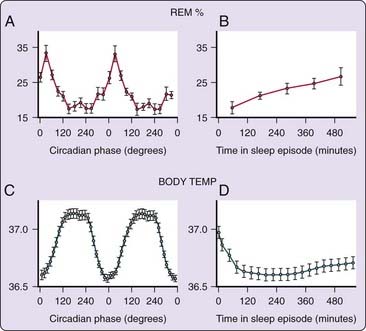
Figure 49-3 Using a forced desynchrony protocol, Dijk and Czeisler44 showed that the sleep-dependent increase in REM propensity (B) parallels a gradual sleep-dependent increase in core body temperature (D) and is distinct from a circadian oscillation in REM propensity (A, double-plotted). The latter tends to vary inversely with body temperature, reaching its acrophase at about 8 AM, roughly 6 hours before onset of the core body temperature plateau (C, double-plotted).
Applying these REM propensity patterns to dreaming, the following sections examine whether across-the-night changes in dreaming may be identified that are characterized by circadian (abrupt, switchlike) and sleep-dependent (gradual, linear) oscillations. Some changes in dream length, content, organization, and memory sources suggest circadian-style changes whereas others suggest sleep-dependent changes. In the case of circadian changes, marked differences are noted between reports from the first third of the night and all later sample points (see reviews in Nielsen 20041 and Nielsen 20052).
Dream Recall and Report Length Changes Across the Night
Some studies reveal gradual across-the-night changes that resemble sleep-dependent effects. There is a lengthening of young adults’ dream reports sampled from early (0 to 2.5 hours) to middle (2.5 to 5 hours) to late (5 to 7.5 hours) night when awakenings are conducted at a constant 4.8 to 5 minutes into REM sleep.23,46 However, for older subjects the increase in length occurs only between middle- and late-night samples. Similarly, when mentation is sampled from the first four REM periods (awakenings all 9 minutes into each stage), a gradual increase across the night is observed for one story structure measure, the number of episodes per story (P < .001; order of means: REM 1 < REM 2 < REM 3 = REM 4) but not another, the number of statements in the event structure (P < .001; order of means: REM 1 < REM 2 = REM 3 = REM 4).47
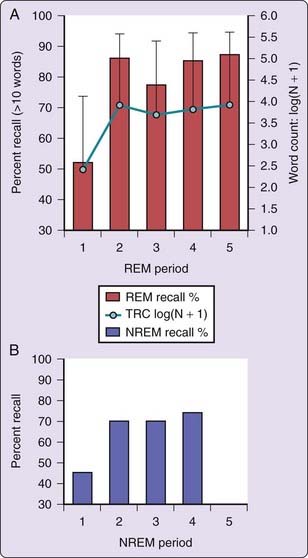
Other findings more consistently suggest circadian switchlike changes similar to those for REM propensity and core body temperature (CBT). Our assessment of 40 subjects (135 reports)48 found that both probability of recall and mean word count were lower for REM 1 than for REM 2 through REM 5 with no difference among the latter (Fig. 49-4, A panel). Because REM awakenings were made progressively later into each REM period (5 minutes into REM 1, 10 minutes into REM 2, 15 minutes into REM 3, 20 minutes into REM 4 and REM 5); however, these findings may be confounded by an ultradian oscillation.
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


