Chapter 21 Autonomic Nervous System
The autonomic nervous system represents the visceral component of the nervous system. It consists of neurones located within both the central nervous system (CNS) and the peripheral nervous system (PNS) and is concerned with control of the internal environment through innervation of secretory glands and with cardiac and smooth muscle. The term ‘autonomic’ is a convenient rather than appropriate title, because the functional autonomy of this part of the nervous system is illusory. Rather, its functions are normally closely integrated with changes in somatic activities, although the anatomical bases for such interactions are not always clear.
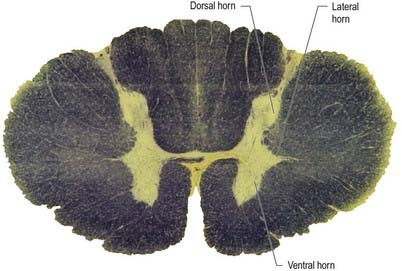
Visceral efferent pathways differ from their somatic equivalents, in that the former are interrupted by peripheral synapses; there is a sequence of at least two neurones between the CNS and the target structure (Fig. 21.1). These are referred to as preganglionic and postganglionic neurones. The somata of preganglionic neurones are located in the visceral efferent nuclei of the brain stem and in the lateral grey columns of the spinal cord. Their axons, which are usually finely myelinated, exit from the CNS in certain cranial and spinal nerves and then pass to peripheral ganglia, where they synapse with the postganglionic neurones. The axons of postganglionic neurones are usually non-myelinated. Postganglionic neurones are more numerous than preganglionic ones; one preganglionic neurone may synapse with 15 to 20 postganglionic neurones, which permits the wide diffusion of many autonomic effects.
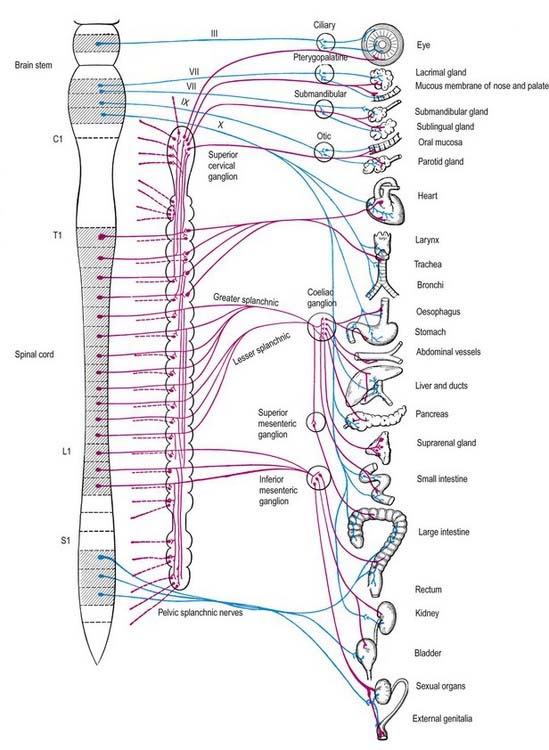
Sympathetic Nervous System
The cell bodies of preganglionic sympathetic neurones are located in the lateral horn of the spinal grey matter of all thoracic segments and the upper two or three lumbar segments (Fig. 21.2). Their axons are myelinated, with diameters of 1.5 to 4 µm. These leave the cord in corresponding ventral nerve roots and pass into the spinal nerves, but they soon leave in white rami communicantes to join the sympathetic trunk (Fig. 21.3). Neurones like those in the lateral grey column exist at other levels of the cord above and below the thoracolumbar outflow, and small numbers of their fibres leave in other ventral roots. Preganglionic sympathetic neurones release acetylcholine as their principal neurotransmitter.

On reaching the sympathetic trunk, preganglionic fibres may behave in one of several ways (see Fig. 21.3). They may synapse with neurones in the nearest ganglion or traverse the nearest ganglion and ascend or descend in the sympathetic chain to end in another ganglion. A preganglionic fibre may terminate in a single ganglion or, through collateral branches, synapse with neurones in several ganglia. Preganglionic fibres may traverse the nearest ganglion, ascend or descend and, without synapsing, emerge in one of the medially directed branches of the sympathetic trunk to synapse in the ganglia of autonomic plexuses (situated mainly in the midline, such as around the coeliac and mesenteric arteries). More than one preganglionic fibre may synapse with a single postganglionic neurone. Uniquely, the suprarenal gland is innervated directly by preganglionic sympathetic neurones that traverse the sympathetic trunk and coeliac ganglion without synapse.
The somata of sympathetic postganglionic neurones are located mostly in ganglia of the sympathetic trunk or ganglia in more peripheral plexuses. Therefore, the axons of postganglionic neurones are generally longer than those of preganglionic neurones; an exception is some of those that innervate pelvic viscera. The axons of ganglionic cells are non-myelinated. They are distributed to target organs in various ways. Those from a ganglion of the sympathetic trunk may return to the spinal nerve of preganglionic origin through a grey ramus communicans, which usually joins the nerve just proximal to the white ramus; they are then distributed through ventral and dorsal spinal rami to blood vessels, sweat glands, hairs and so forth in their zone of supply. Segmental areas vary in extent and overlap considerably. The extent of innervation of different effector systems (e.g. vasomotor, sudomotor) by a particular nerve may not be the same. Alternatively, postganglionic fibres may pass in a medial branch of a ganglion directly to particular viscera, or they may innervate adjacent blood vessels or pass along them externally to their peripheral distribution. They may ascend or descend before leaving the sympathetic trunk. Many fibres are distributed along arteries and ducts as plexuses to distant effectors.
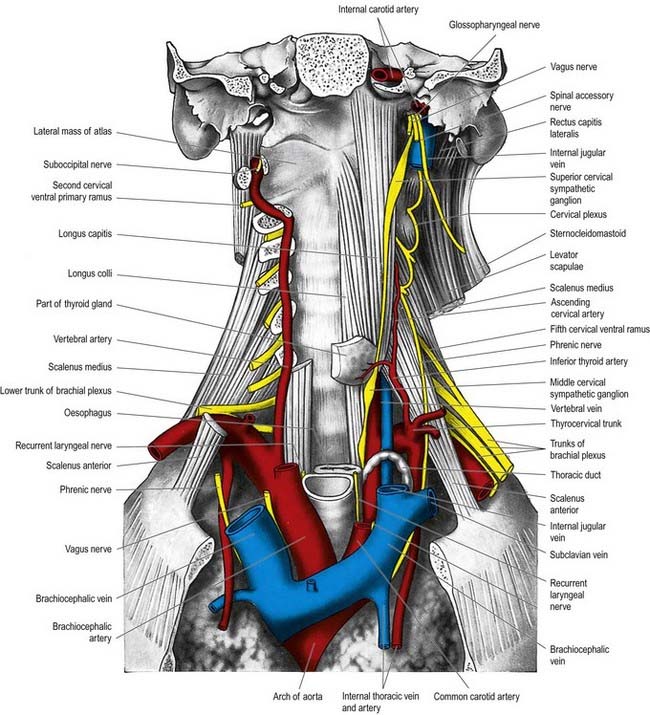
Cervical Sympathetic Trunk
The cervical sympathetic trunk (Figs 21.4, 21.5) lies on the prevertebral fascia behind the carotid sheath and contains three interconnected ganglia: the superior, middle and inferior (stellate or cervicothoracic). However, there may occasionally be two or four ganglia. The cervical sympathetic ganglia send grey rami communicantes to all the cervical spinal nerves but receive no white rami communicantes from them. Their spinal preganglionic fibres emerge in the white rami communicantes of the upper five thoracic spinal nerves (mainly the upper three) and ascend in the sympathetic trunk to synapse in the cervical ganglia. In their course, the grey rami communicantes may pierce longus capitis or scalenus anterior.
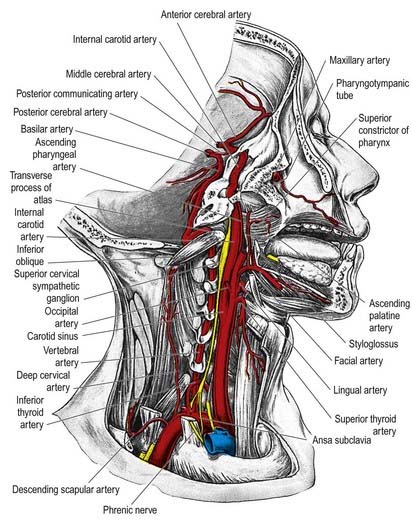
Superior Cervical Ganglion
The superior cervical ganglion is the largest of the three ganglia. It lies on the transverse processes of the second and third cervical vertebrae and is probably formed from four fused ganglia, judging by its grey rami to C1–4. The internal carotid artery within the carotid sheath is anterior, and longus capitis is posterior (see Fig. 21.4). The lower end of the ganglion is united by a connecting trunk to the middle cervical ganglion. Postganglionic branches are distributed in the internal carotid nerve, which ascends with the internal carotid artery into the carotid canal to enter the cranial cavity, and in lateral, medial and anterior branches. They supply vasoconstrictor and sudomotor nerves to the face and neck, dilator pupillae and smooth muscle in the eyelids and orbitalis.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


